Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists
Proceedings of the Integrated Ocean Drilling Program Volume 320/321
publications.iodp.org

https://doi.org/10.2204/iodp.proc.320321.221.2020
Data report: middle Miocene planktonic foraminifers from the eastern equatorial Pacific, IODP Expedition 321 Site U13381
Takuma Suzuki,2 Hiroki Hayashi,2 and Kyoko Idemitsu2
Keywords: Integrated Ocean Drilling Program, IODP, JOIDES Resolution, Expedition 321, Site U1338, equatorial Pacific, middle Miocene, planktonic foraminifers
MS 320321-221: Received 30 January 2019 · Accepted 16 April 2020 · Published 24 June 2020
Abstract
Middle Miocene planktonic foraminiferal assemblages are reported using core samples from Site U1338, which was drilled during Integrated Ocean Drilling Program (IODP) Expedition 321 in the eastern equatorial Pacific. A total of 63 species belonging to 20 genera were classified from 98 samples. In addition, we reexamined biostratigraphic events of the Fohsella lineage following recently proposed taxonomic criteria. We recognized 10 biohorizons of the Fohsella lineage. Among them, the numerical ages of eight events are restricted by orbitally tuned ages.
Introduction
The Pacific Equatorial Age Transect (PEAT), which included Integrated Ocean Drilling Program (IODP) Expeditions 320 and 321, was designed to obtain sediments throughout the Cenozoic. From 5 March to 22 June 2009, Sites U1331–U1338 were drilled in the eastern equatorial Pacific (Figure F1). At Site U1338 (2°30.469ʹN, 117°58.178ʹE; 4200 m water depth), a continuous succession of sediments from the latest Oligocene to the present were obtained (Pälike et al., 2010).
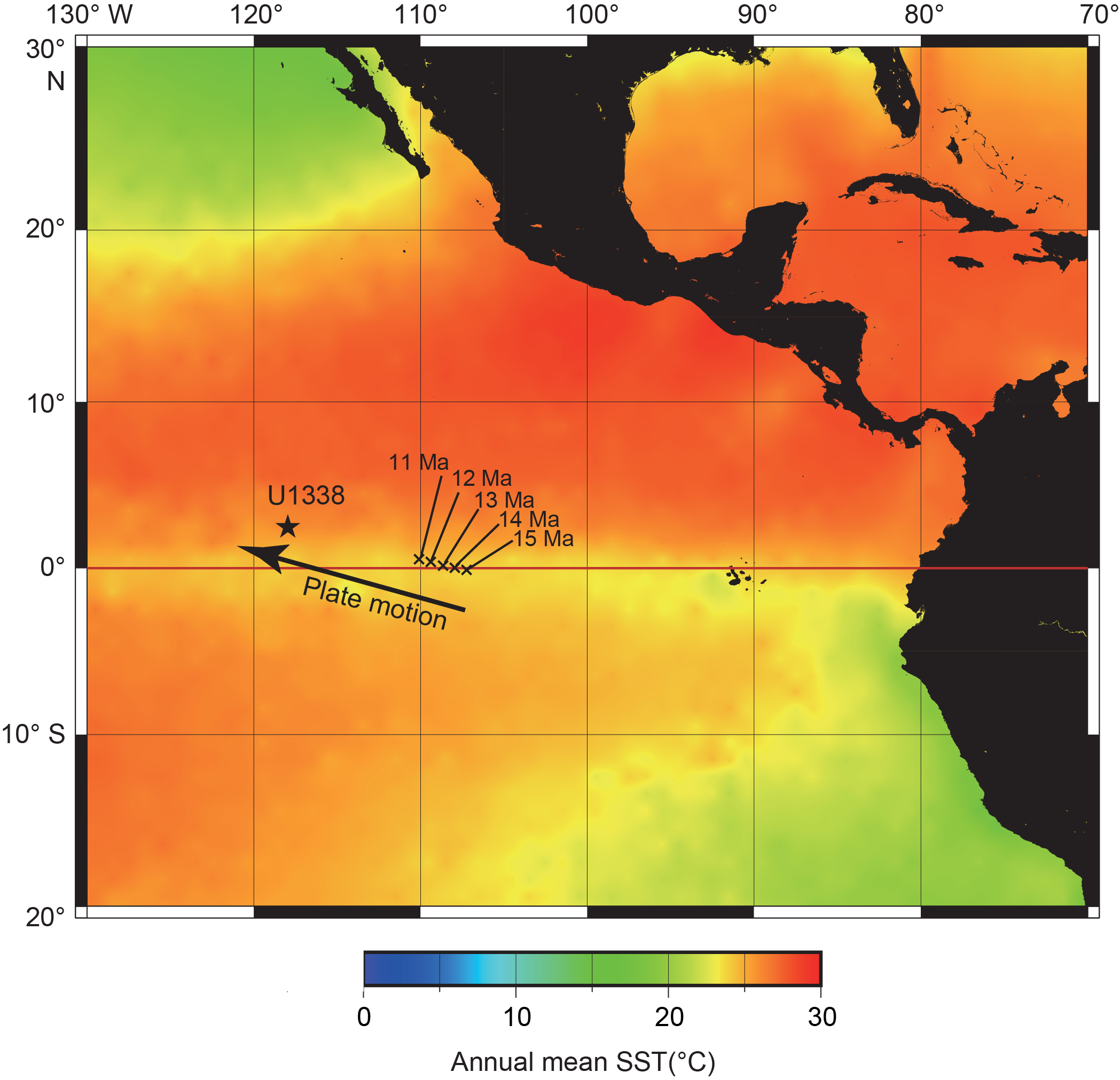
Figure F1. Location map.
The shipboard composite depth scale for Site U1338 (Pälike et al., 2010) was revised by Wilkens et al. (2013). On the new depth scale, Backman et al. (2016) proposed a revised biomagnetostratigraphic age model for Site U1338. For the interval from 16 to 13 Ma at the site, Holbourn et al. (2014) made an orbitally tuned age model by integrating high-resolution benthic foraminiferal stable isotope data and X-ray fluorescence scanner–derived biogenic silica and carbonate accumulation. The integration of such recent works enables us to determine the precise numerical ages of biohorizons.
Hayashi et al. (2013) reported the middle Miocene to Pleistocene planktonic foraminiferal biostratigraphy at Site U1338. They investigated Cores 321-U1338B-1H through 40H at a sampling interval of one per section and recognized 60 biohorizons. The assemblage data have not yet been published. Thus, the first purpose of this data report is to describe the assemblage data of Hayashi et al. (2013) for the middle Miocene interval.
After the completion of the biostratigraphic work by Hayashi et al. (2013), a new classification for the Fohsella lineage was proposed by Si and Berggren (2017). They reconsidered the type specimens and specimens from continuous deep-sea sediments (Ocean Drilling Program Site 806 at Ontong Java Plateau) and recognized two branched evolutionary transitions: Fohsella peripheroacuta–Fohsella praefohsi–Fohsella lobata and F. peripheroacuta–F. “praefohsi”–Fohsella fohsi–Fohsella robusta. The second purpose of this report is to reexamine the Fohsella lineage biohorizons of Hayashi et al. (2013) by applying Si and Berggren’s (2017) classification.
Methods and materials
A total of 104 samples were collected from Cores 321-U1338B-25H through 40H (228.9–371.3 m core composite depth below seafloor, Method B [CCSF-B]) at an average depth interval of 1.5 m (one sample per section). These samples correspond to the lower samples of Hayashi et al. (2013) but were newly treated in the present study. The depth scale references are based on Wilkens et al. (2013).
Sediment samples of 1–5 g dry weight were disaggregated using a 3% hydrogen peroxide solution. After the samples had been macerated, each sample was wet sieved through a 63 µm screen. The dried samples were then divided into suitable volumes using a sample splitter, yielding approximately 200 planktonic foraminiferal specimens. Planktonic foraminiferal specimens larger than 125 µm were observed under a binocular microscope. We conducted classifications of each counting split to identify all planktonic foraminifers. We then made additional observations of the washed residue for important horizons to detect index species. Species detected by additional observations are recorded as “+” in the occurrence table (Table T1). Scanning electron photomicrographs of Fohsella species were obtained with a JCM-5000 (JEOL Co. Ltd., Japan).
Preservation of each sample is recorded in Table T1 using the following criteria (see Expedition 320/321 Scientists, 2010a):
- VG = very good (no evidence of overgrowth, dissolution, or abrasion).
- G = good (little evidence of overgrowth, dissolution, or abrasion).
- M = moderate (calcite overgrowth, dissolution, or abrasion is common but minor).
- P = poor (substantial overgrowth, dissolution, or fragmentation).
The taxonomic names and biochronology in this study generally follow Hayashi et al. (2013), partly modified to agree with recent taxonomic studies (Fox and Wade, 2013; Spezzaferri et al., 2015; Si and Berggren, 2017; Poole and Wade, 2019).
The age model for the present study was established by the astronomically tuned age model of Holbourn et al. (2014) before 12.72 Ma and the shipboard biomagnetostratigraphic age model of Expedition 320/321 Scientists (2010b) revised by Backman et al. (2016) after 12.72 Ma (Figure F2). To calculate the planktonic foraminiferal flux (number/cm2/ky) for each sample, we assumed the dry density of each sediment using the average of the two nearest horizons (above and below the sample) of the shipboard moisture and density measurements (Expedition 320/321 Scientists, 2010b). The planktonic foraminiferal flux and the planktonic foraminifer ratio (P/T) are shown in Table T1.
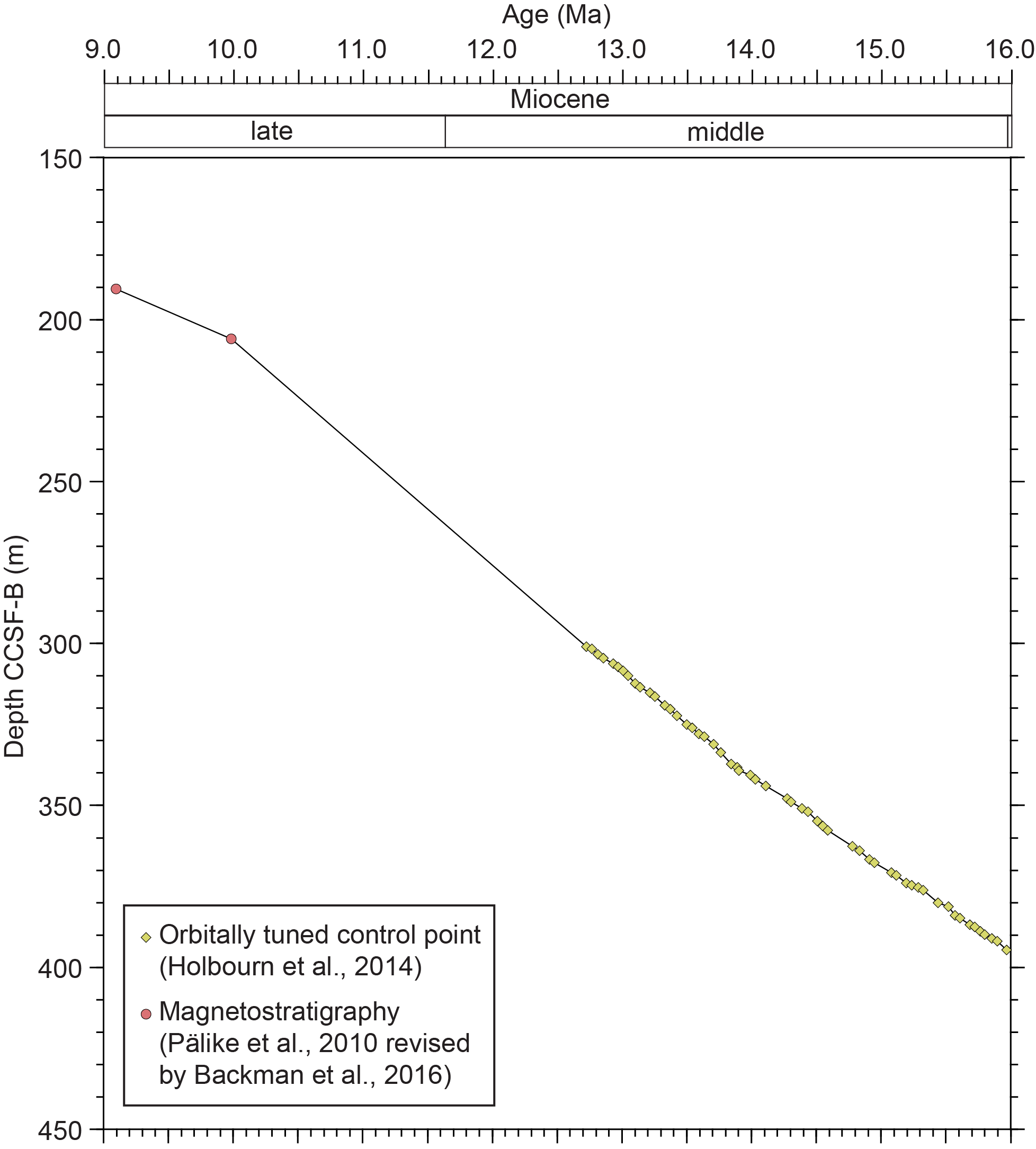
Figure F2. Age-depth model.
Results
A total of 16,323 planktonic foraminiferal individuals were collected from 98 samples and classified into 63 species belonging to 20 genera. Preservation of the tests was generally good. However, in the lower part of the studied interval, preservation became relatively poor owing to the fragmentation of the tests and dissolution on the surface of the test.
In the relative abundance of all samples, Paragloborotalia siakensis sinistral form had the highest occurrence (37.5%) (Figure F3). The top 10 species other than P. siakensis were Globigerinita glutinata (12.4%), Paragloborotalia continuosa sinistral form (8.2%), Dentoglobigerina venezuelana (6.0%), P. siakensis dextral form (3.8%), Neogloboquadrina cf. pseudopachyderma sinistral form (3.4%), Sphaeroidinellopsis seminulina (2.8%), Globorotalia praemenardii (2.6%), Dentoglobigerina altispira altispira (2.2%), Globigerinita uvula (2.0%), and Neogloboquadrina praeatlantica sinistral form (1.7%). Species belonging to the Fohsella linages were commonly observed (2.6%) below Sample 321-U1338B-27H-6, 38–40 cm, and the highest occurrence of 19.6% was in Sample 35H-2, 37–29 cm. The mean planktonic foraminiferal flux was 3494 (number/cm2/ky). The maximum value was 71,806 in Sample 40H-2, 37–39 cm, and the minimum value was 0 in Sample 25H-6, 38–40 cm.
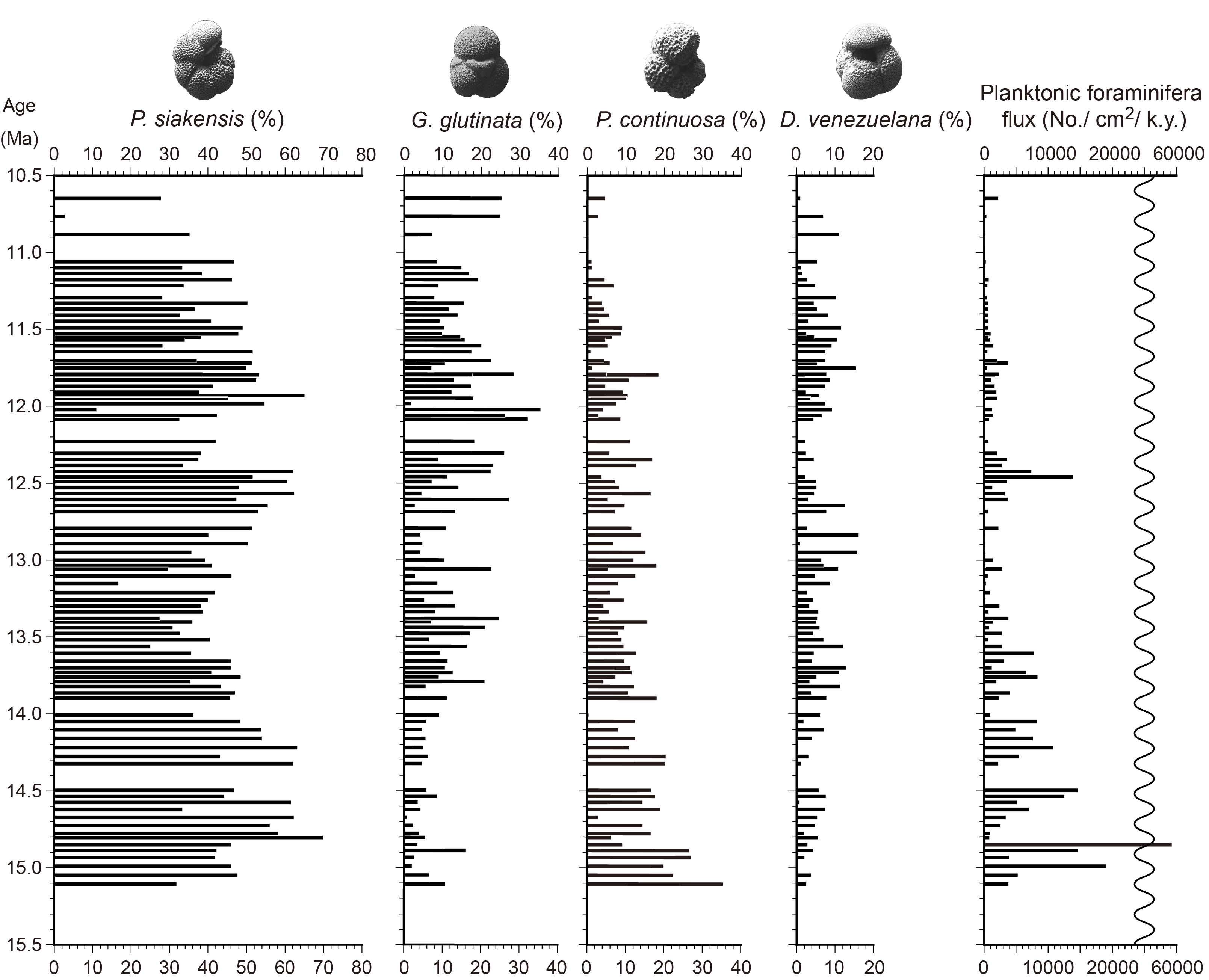
Figure F3. Relative abundance.
The biohorizons of the study interval were reported by Hayashi et al. (2013). It should be noted that the taxonomic criteria for the important evolutionary lineage Fohsella spp. have been controversial for a long time, as reviewed by Wade et al. (2011). Recently, the classification of the lineage was revised by Si and Berggren (2017) using type specimens together with deep-sea samples. We reexamined our Fohsella specimens using the Si and Berggren (2017) classification (Figure F4). The newly proposed classification is highly adaptable to the present sequence. A total of 10 biohorizons of Fohsella chronospecies were recognized in the quality criteria (A–C) of Hayashi et al. (2013). The numerical ages of the 10 biohorizons at Site U1338 were calculated by using the revised age-depth model (Table T2).
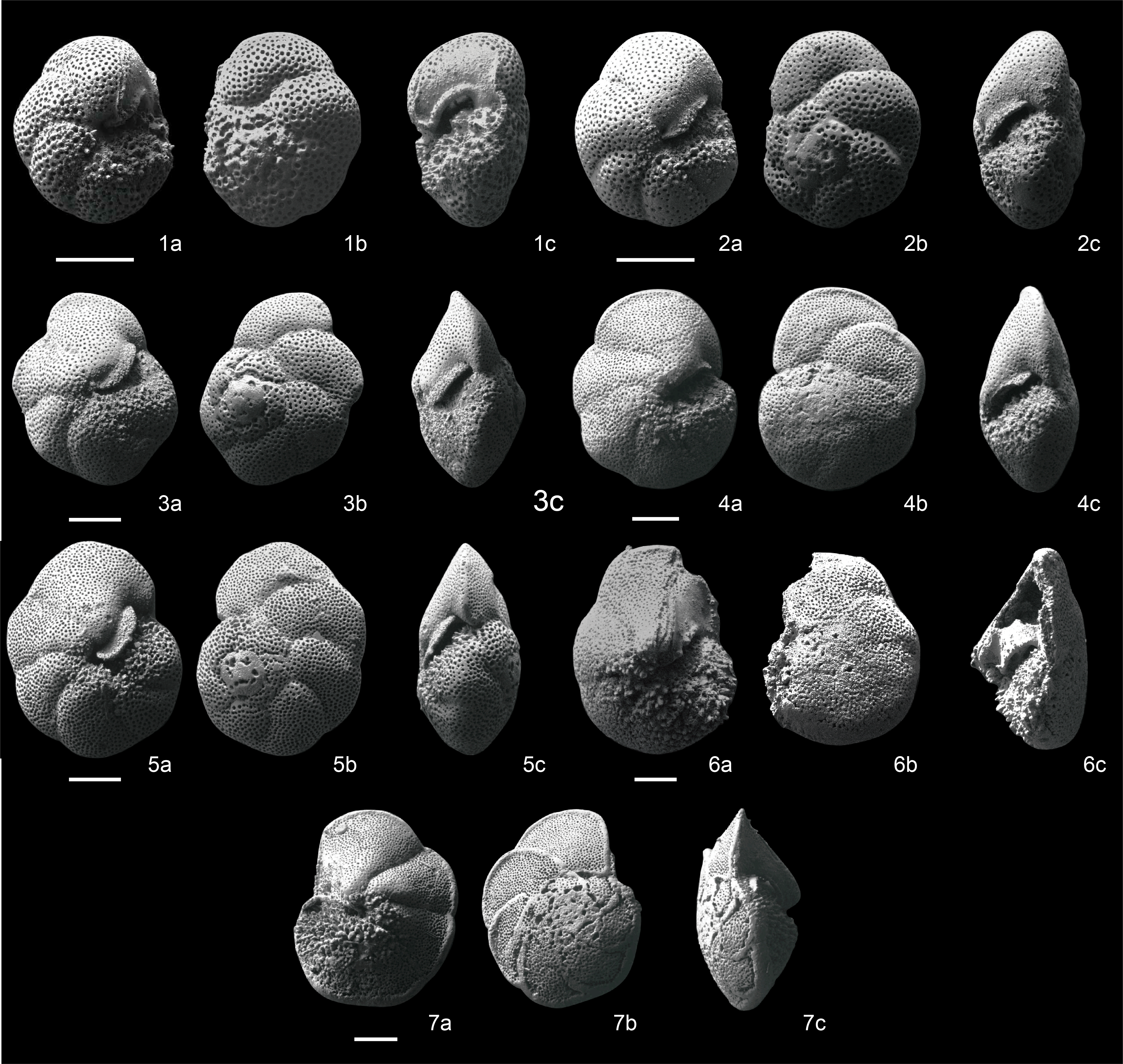
Figure F4. Fohsella species.
Among the 10 biohorizons (Figure F5), 3 biohorizons are newly distinguished in this study: the base occurrence of F. praefohsi, the top occurrence (TO) of Fohsella peripheroronda, and the top common occurrence of Fohsella spp. Another three biohorizons (2, 7, and 9 in Table T2) were reported by Hayashi et al. (2013), but their stratigraphic positions were revised by the taxonomic reexamination. Based on comparison with the Geologic Time Scale 2012 (GTS2012; Hilgen et al., 2012), the numerical ages calculated in the present study are slightly older than those of GTS2012, except for the TO of F. peripheroronda, which is significantly younger than GTS2012. This biohorizon has been regarded as latitudinal diachrony in previous studies (e.g., Takayanagi et al., 1984; Spencer-Cervato et al., 1994). This revised data set of biohorizons will provide a good framework to establish the standard timescale for the eastern equatorial Pacific region.
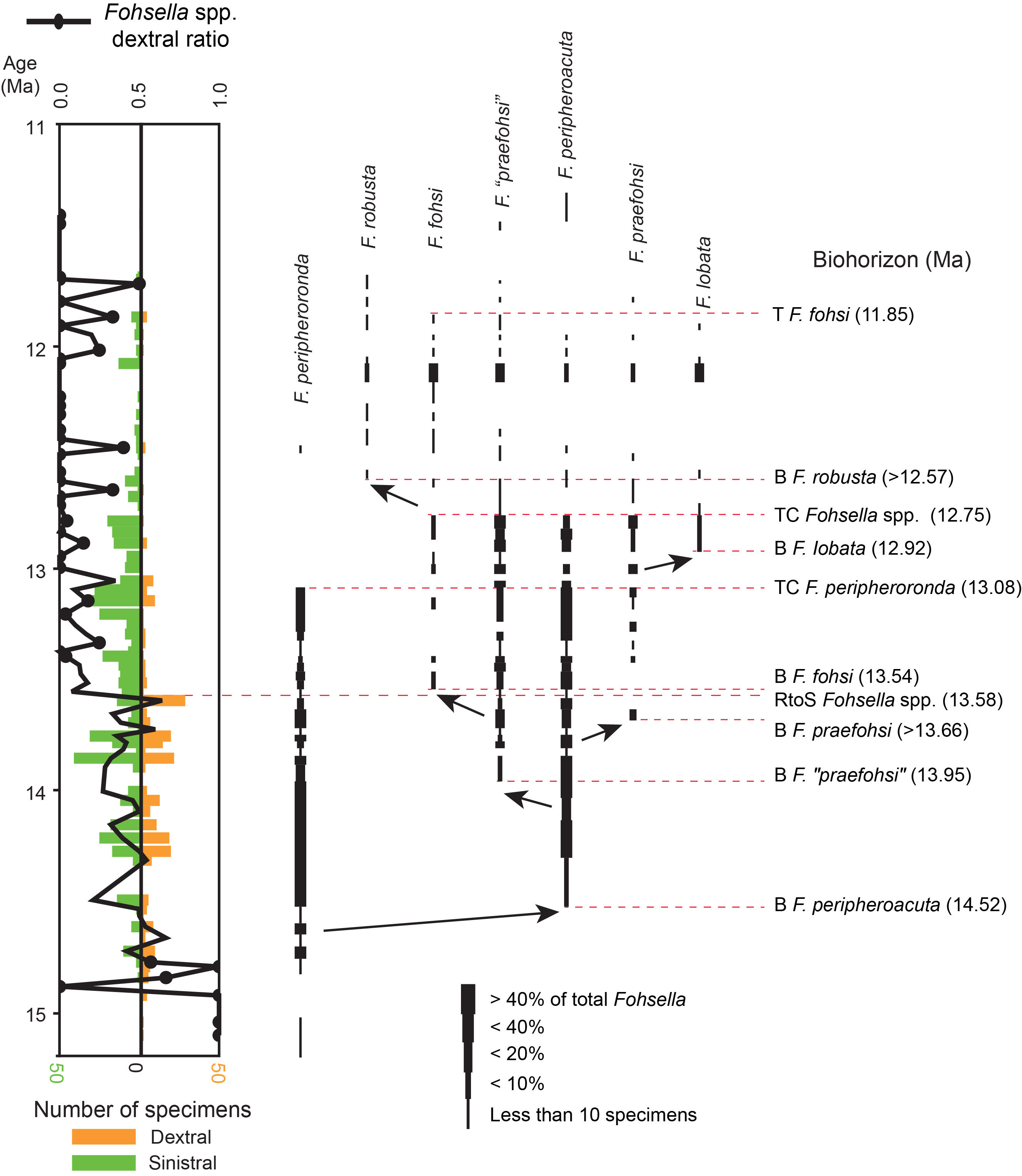
Figure F5. Temporal biohorizon distribution.
Acknowledgments
This research used samples provided by the Integrated Ocean Drilling Program (IODP). The study was funded by Grants-in-Aid 23740377 and 15K05311 from the Japan Society for the Promotion of Science. Figures F1, F2, F3, and F5 were produced by GMT software developed by Wessel and Smith (1991).
References
Akers, W.H., 1955. Some planktonic foraminifera of the American gulf coast and suggested correlations with the Caribbean Tertiary. Journal of Paleontology, 29(4):647–664. https://www.jstor.org/stable/1300351
Backman, J., Baldauf, J.G., Ciummelli, M., and Raffi, I., 2016. Data report: a revised biomagnetostratigraphic age model for Site U1338, IODP Expedition 320/321. In Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists, Proceedings of the Integrated Ocean Drilling Program, 320/321: Tokyo (Integrated Ocean Drilling Program Management International, Inc.). https://doi.org/10.2204/iodp.proc.320321.219.2016
Banner, F.T., and Blow, W.H., 1960. Some primary types of species belonging to the superfamily Globigerinaceae. Contributions from the Cushman Foundation for Foraminiferal Research, 11(1):1–41.
Berggren, W.A., 1972. Cenozoic biostratigraphy and paleobiogeography of North Atlantic. In Laughton, A.S., Berggren, W.A., et al., Initial Reports of the Deep Sea Drilling Project, 12: Washington (US Government Printing Office), 965–1001. https://doi.org/10.2973/dsdp.proc.12.114.1972
Bermúdez, P.J., 1949. Tertiary smaller foraminifera of the Dominican Republic. Special Publication - Cushman Foundation for Foraminiferal Research, 25.
Bermúdez, P.J., 1961. Contribuciön al estudio de las Globigerinidae de la regiön Caribe-Antillana: Paleocene-Recente. Mem. III Congr. Geol. Venezolano. Soc. Geol. Venez., Bol. Geol., Publ. Espec., 3:1119–1393.
Blow, W.H., 1956. Origin and evolution of the foraminiferal genus Orbulina d’Orbigny. Micropaleontology, 2(1):57–70. https://doi.org/10.2307/1484492
Blow, W.H., 1959. Age, correlation and biostratigraphy of the upper Tocuyo (San Lorenzo) and Pozón formations, eastern Falcon, Venezuela. Bulletins of American Paleontology, 39(178):59–251.
Blow, W.H., and Banner, F.T., 1966. The morphology, taxonomy and biostratigraphy of Globorotalia barisanensis LeRoy, Globorotalia fohsi Cushman and Ellisor, and related taxa. Micropaleontology, 12(3):286–302. https://doi.org/10.2307/1484548
Bolli, H.M., 1950. The direction of coiling in the evolution of some Globorotaliidae. Contributions from the Cushman Foundation for Foraminiferal Research, 1:82–89.
Bolli, H.M., 1957. Planktonic foraminifera from the Oligocene-Miocene Cipero and Lengua formations of Trinidad, B.W.I. In Loeblich, A.R., Jr., Tappan, H., Beckmann, J.P., Bolli, H.M., Gallitelli, E.M., and Troelsen, J.C. (Eds.), Studies in Foraminifera. Bulletin - United States National Museum, 215:97–123.
Bolli, H.M., Loeblich, A.R., Jr., and Tappan, H., 1957. Planktonic foraminiferal families Hantkeninidae, Orbulinidae, Globorotaliidae, and Globotruncanidae. Bulletin - United States National Museum, 215:3–50.
Brady, H.B., 1882. Report on the foraminifera. In Tizard, L., and Murray, J. (Eds.), Exploration of the Faröe Channel During the Summer of 1880, in Her Majesty’s Ship Knight Errant, with Subsidiary Reports. Proceedings of the Royal Society of Edinburgh, 11:708–717.
Brönnimann, P., 1951. The genus Orbulina d’Orbigny in the Oligo–Miocene of Trinidad, B.W.I. Contributions from the Cushman Foundation for Foraminiferal Research, 2(4):131–138.
Chapman, F., Parr, W.J., and Collins, A.C., 1934. Tertiary foraminifera of Victoria, Australia: the Balcombian deposits of Port Philip (Pt. III). Zoological Journal of the Linnean Society, 38(262):553–577. https://doi.org/10.1111/j.1096-3642.1934.tb00996.x
Cita, M.B., Premoli Silva, I., and Rossi, R., 1965. Foraminiferi planctonici del Tortoniano-tipo. Rivista Italiana di Paleontologia e Stratigrafia, 71:217–308.
Cushman, J.A., and Ellisor, A.C., 1939. New species of foraminifera from the Oligocene and Miocene. Contributions from the Cushman Laboratory for Foraminiferal Research, 15:1–14.
Cushman, J.A., and Jarvis, P.W., 1936. Three new foraminifera from the Miocene Bowden marl of Jamaica. Contributions from the Cushman Foundation for Foraminiferal Research, 12(1):3–5.
Cushman, J.A., and Stainforth, R.M., 1945. The foraminifera of the Cipero marl formation of Trinidad, British West Indies. Special Publication - Cushman Laboratory for Foraminiferal Research, 14:1–75.
d’Orbigny, A.D., 1839. Foraminifères. In de la Sagra, R. (Ed.), Histoire Physique, Politique et Naturelle de Lîle de Cuba: Paris (Arthus Bertrand), 8:1–224.
d’Orbigny, A., 1846. Foraminifères Fossiles du Bassin Tertiaire de Vienne (Autriche): Paris (Gide et Comp).
Egger, J.G., 1893. Foraminiferen aus Meeresgrundproben Gelothet von 1874 bis 1876 von S. M. Sch. Gazelle. Abhandlungen, Bayerische Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse, 18:193–458.
Ehrenberg, C.G., 1861. Elemente des tiefen Meeresgrundes in Mexikanischen Golfstrome bei Florida: Ueber die Tiefgrund-Verhältnisse des Oceans am Eingang der Davisstrasse und bei Island. Monatsberichte der Königlichen Preussische Akademie des Wissenschaften zu Berlin, 222–240, 275–315.
Expedition 320/321 Scientists, 2010a. Methods. In Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists. Proceedings of the Integrated Ocean Drilling Program, 320/321: Tokyo (Integrated Ocean Drilling Program Management International, Inc.). https://doi.org/10.2204/iodp.proc.320321.102.2010
Expedition 320/321 Scientists, 2010b. Site U1338. In Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists, Proceedings of the Integrated Ocean Drilling Program, 320/321: Tokyo (Integrated Ocean Drilling Program Management International, Inc.). https://doi.org/10.2204/iodp.proc.320321.110.2010
Finlay, H.J., 1939. New Zealand foraminifera: key species in stratigraphy, No. 3. Transactions of the Royal Society of New Zealand, 69(3):309–329.
Foresi, L.M., Iaccarino, S.M., and Salvatorini, G., 2002. Neogloboquadrina atlantica praeatlantica, new subspecies from late middle Miocene. Rivista Italiana di Paleontologia e Stratigrafia, 108(2):325–336.
https://doi.org/10.13130/2039-4942/5479
Fox, L., and Wade, B.S., 2013. Systematic taxonomy of early–middle Miocene planktonic foraminifera from the equatorial Pacific Ocean: Integrated Ocean Drilling Program, Site U1338. Journal of Foraminiferal Research, 44(4):373–404. https://doi.org/10.2113/gsjfr.43.4.374
Giannelli, L., and Salvatorini, G., 1976. Due nuove specie di foraminiferi planctonici del Miocene. Bollettino della Società Paleontologica Italiana, 15:167–173.
Hayashi, H., Idemitsu, K., Wade, B.S., Idehara, Y., Kimoto, K., Nishi, H., and Matsui, H., 2013. Middle Miocene to Pleistocene planktonic foraminiferal biostratigraphy in the eastern equatorial Pacific Ocean. Paleontological Research, 17(1):91–109. https://doi.org/10.2517/1342-8144-17.1.91
Hedberg, H.D., 1937. Foraminifera of the middle Tertiary Carapita formation of northeastern Venezuela. Journal of Paleontology, 11(8):661–697. https://www.jstor.org/stable/1298440
Hilgen, F.J., Lourens, L.J., and Van Dam, J.A., 2012. The Neogene period. With contributions by A.G. Beu, A.F. Boyes, R.A. Cooper, W. Krijgsman, J.G. Ogg, W.E. Piller, and D.S. Wilson. In Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. (Eds.), The Geologic Time Scale: Oxford, United Kingdom (Elsevier), 923–978. https://doi.org/10.1016/B978-0-444-59425-9.00029-9
Holbourn, A., Kuhnt, W., Lyle, M., Schneider, L., Romero, O., and Anderson, N., 2014. Middle Miocene climate cooling linked to intensification of eastern equatorial Pacific upwelling. Geology, 42(1):19–22. https://doi.org/10.1130/G34890.1
Jenkins, D.G., 1960. Planktonic foraminifera from the Lakes Entrance oil shaft, Victoria, Australia. Micropaleontology, 6(4):345–371. https://doi.org/10.2307/1484217
Kennett, J.P., and Srinivasan, M.S., 1983. Neogene Planktonic Foraminifera: A Phylogenetic Atlas: Stroudsburg, PA (Hutchinson Ross).
Koppers, A.A.P., Morgan, J.P., Morgan, J.W., and Staudigel, H., 2001. Testing the fixed hotspot hypothesis using 40Ar/39Ar age progressions along seamount trails. Earth and Planetary Science Letters, 185(3–4):237–252. https://doi.org/10.1016/S0012-821X(00)00387-3
LeRoy, L.W., 1939. Some small foraminifera, ostracoda and otoliths from the Neogene (“Miocene”) of the Rokantapanoeli area, Central Sumatra. Natuurwetenschappelijk Tijdschrift voor Nederlandsch Indie, 99:215–296.
Natland, M.L., 1938. New species of Foraminifera from off the West Coast of North America and from the later Tertiary of the Los Angeles basin. Bulletin of the Scripps Institution of Oceanography, Technical Series, 4(5):137–164.
Pälike, H., Nishi, H., Lyle, M., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists, 2010. Expedition 320/321 summary. In Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists, Proceedings of the Integrated Ocean Drilling Program, 320/321: Tokyo (Integrated Ocean Drilling Program Management International, Inc.). https://doi.org/10.2204/iodp.proc.320321.101.2010
Poole, C.R., and Wade, B.S., 2019. Systematic taxonomy of the Trilobatus sacculifer plexus and descendant Globigerinoidesella fistulosa (planktonic foraminifera). Journal of Systematic Paleontology, 17(23):1989–2030. https://doi.org/10.1080/14772019.2019.1578831
Reuss, A.E., 1850. Neue Foraminiferen aus den Schichten des Oesterreichischen Tertiärbeckens. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe, Denkschrift, 1:365–390.
Saito, T., 1963. Miocene planktonic foraminifera from Honshu, Japan. The Science Report of the Tohoku University, 2nd Series, 35(2):123–209.
Saito, T., and Maiya, S., 1973. Planktonic foraminifera of the Nishikurosawa Formation, northeast Honshu, Japan. Transactions and Proceedings of the Paleontological Society of Japan, 1973(91):113–125. https://doi.org/10.14825/prpsj1951.1973.91_113
Schwager, C., 1866. Fossile Foraminiferen von Kar Nikobar. Novara Expedition, 1857–1859, Wein, Geol. Theile, 2:187–268.
Si, W., and Berggren, W.A., 2017. Taxonomy, stratigraphy and phylogeny of the middle Miocene Fohsella lineage: geometric morphometric evidence. Journal of Foraminiferal Research, 47(4):310–324. https://doi.org/10.2113/gsjfr.47.4.310
Spencer-Cervato, C., Thierstein, H.R., Lazarus, D.B., and Beckmann, J.-P., 1994. How synchronous are Neogene marine plankton events? Paleoceanography and Paleoclimatology, 9(5):739–763. https://doi.org/10.1029/94PA01456
Spezzaferri, S., Kucera, M., Pearson, P.N., Wade, B.S., Rappo, S., Poole, C.R., Morard, R., and Stalder, C., 2015. Fossil and genetic evidence for the polyphyletic nature of the planktonic foraminifera “Globigerinoides,” and description of the new genus Trilobatus. PLoS One, 10(5):e0128108. https://doi.org/10.1371/journal.pone.0128108
Subbotina, N.N., Pishvanova, L.S., and Ivanova, L.V., 1960. Stratigrafiya oligotsenovikh i miotsenovikh otlozheniy Predkarpatya po foraminiferam. Mikrofauna SSSR, 11. Trudi VNIGRI, 153:5–127.
Takayanagi, Y., Oda, M., Hasegawas, S., Honda, N., Maruyama, T., and Funayama, M., 1984. Some middle Miocene planktonic microfossil datum planes in northern Honshu, Japan: their paleoceanographic implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 46(1–3):71–84. https://doi.org/10.1016/0031-0182(84)90026-9
Takayanagi, Y., and Saito, T., 1962. Planktonic foraminifera from the Nobori Formation, Shikoku, Japan. Science Reports of the Tohoku University, 2nd Series, 5:67–106.
Takayanagi, Y., Takayama, T., Sakai, T., Oda, M., and Kitazato, H., 1976. Microbiostratigraphy of some middle Miocene sequences in northern Japan. In Asano, K., Takayanagi, Y., and Saito, T. (Eds.), Progress in Micropaleontology: New York (Micropaleontology Press), 356–381.
Todd, R., 1957. Smaller foraminifera. In Johnson, J.H., Bramlette, M.N., Reidel, W.R., Todd, R., Cole, W.S., and Cooke, C.W. (Eds.), Geology of Saipan, Mariana Islands. Part 3, Paleontology. U.S. Geological Survey Professional Paper, 280(E–J):265–320.
Wade, B.S., Pearson, P.N., Berggren, W.A., and Pälike, H., 2011. Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth-Science Reviews, 104(1–3):111–142. https://doi.org/10.1016/j.earscirev.2010.09.003
Walters, R., 1965. The Globorotalia zealandica and G. miozea lineages. New Zealand Journal of Geology and Geophysics, 8(1):109–127. https://doi.org/10.1080/00288306.1965.10422135
Wessel, P., and Smith W.H.F., 1991. Free software helps map and display data. Eos, Transactions of the American Geophysical Union, 72(41):441–448. https://doi.org/10.1029/90EO00319
Wilkens, R.H., Dickens, G.R., Tian, J., Backman, J., and the Expedition 320/321 Scientists, 2013. Data report: revised composite depth scales for Sites U1336, U1337, and U1338. In Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists, Proceedings of the Integrated Ocean Drilling Program, 320/321: Tokyo (Integrated Ocean Drilling Program Management International, Inc.). https://doi.org/10.2204/iodp.proc.320321.209.2013
Appendix
Faunal references
Catapsydrax stainforthi Bolli et al., 1957, p. 37, pl. 7, figs. 11a–11c.
Catapsydrax unicavus Bolli et al., 1957, p. 37, pl. 7, figs. 9a–9c.
Clavatorella bermudezi (Bolli), 1957, p. 112, pl. 25, figs. 1a–1c.
Dentoglobigerina altispira altispira (Cushman and Jarvis), 1936, p. 5, pl. 1, figs. 13a–13c.
Dentoglobigerina altispira globosa (Bolli), 1957, p. 111, pl. 24, figs. 9a–10c.
Dentoglobigerina baroemoenensis (LeRoy), 1939, p. 263, pl. 6, figs. 1–2.
Dentoglobigerina venezuelana (Hedberg), 1937, p. 681, pl. 92, figs. 72b.
Fohsella fohsi (Cushman and Ellisor), 1939, p. 12, pl. 2, figs. 6a–6c.
Fohsella lobata (Bermúdez), 1949, p. 286, pl. 22, figs. 15–17.
Fohsella peripheroacuta (Blow and Banner), 1966, p. 294, pl. 1, figs. 2a–2c.
Fohsella peripheroronda (Blow and Banner), 1966, p. 294. pl. 1, figs. 1a–1c.
Fohsella praefohsi (Blow and Banner), 1966, p. 295, pl. 1, figs. 6–7, 10–11.
Fohsella “praefohsi” (Blow and Banner), 1966 sensu Si and Berggren (2017), Kennett and Srinivasan, 1983, p. 98, pl. 22, figs. 7–9.
Fohsella robusta (Bolli), 1950, p. 84, 89, pl. 15, figs. 3a–3c.
Globigerina angustiumbilicata Bolli, 1957, p. 109, pl. 22, figs. 12a–13c.
Globigerina falconensis Blow, 1959, p. 177, pl. 9, figs. 40a–40c, 41.
Globigerina praebulloides Blow, 1959, p. 180, pl. 8, figs. 47a–47c; pl. 9, fig. 48.
Globigerinella obesa (Bolli), 1957, p. 119, pl. 29, figs. 2a, 3.
Globigerinita glutinata (Egger), 1893, p. 371, pl. 13, figs. 19–21.
Globigerinita uvula (Ehrenberg), 1861, pl. 2, figs. 24–25.
Globigerinoides bollii Blow, 1959, p. 189, pl. 10, figs. 65a–65c.
Globigerinoides immaturus LeRoy, 1939, p. 263, pl. 3, figs. 19–21.
Globigerinoides quadrilobatus (d’Orbigny), 1846, p. 164, pl. 9, figs. 7–10.
Globigerinoides subquadratus Brönnimann, 1954, p. 680, pl. 1, figs. 8a–8c.
Globigerinoides trilobus (Reuss), 1850, p. 374, pl. 447, figs. 11a–11c.
Globoconella conoidea (Walters), 1965, p. 124, figs. 8i–8m.
Globoconella miozea Finlay, 1939, p. 326, pl. 29, figs. 159–161.
Globoquadrina dehiscens (Chapman et al.), 1934, p. 569, pl. 11, figs. 36a–36c.
Globorotalia adamantae Saito, 1963, p. 173, pl. 54, figs. 4a–5c.
Globorotalia archeomenardii Bolli, 1957, p. 119, pl. 28, figs. 11a–11c.
Globorotalia birnageae Blow, 1959, p. 210, pl. 17, figs. 108a–108c.
Globorotalia bykovae (Aisenstat), in Subbotina et al. (1960), p. 69, pl. 13, figs. 7a–8b.
Globorotalia ichinosekiensis Takayanagi et al. (1976), p. 374, pl. 1, figs. 1a–1c.
Globorotalia lenguaensis Bolli, 1957, p. 120, pl. 29, figs. 5a–5c.
Globorotalia panda Jenkins, 1960, p. 364, pl. 4, figs. 10a–10c.
Globorotalia praemenardii Cushman and Stainforth, 1945, p. 70, pl. 13, figs. 14a–14c.
Globorotalia praescitula Blow, 1959, p. 221, pl. 19, figs. 128a–128c.
Globorotalia quinifalcata Saito and Maiya, 1973, p. 119, pl. 18, figs. 5a–5c, 6a–6c.
Globorotalia scitula (Brady), 1882, p. 716; Banner and Blow, 1960, pt. 1, p. 27, pl. 5, fig. 5.
Globorotaloides falconarae Giannelli and Salvatorini, 1976, p. 170, pl. 2.
Globorotaloides hexagona (Natland), 1938, p. 149, pl. 7, figs. 1a–1c.
Globorotaloides variabilis Bolli, 1957, p. 117, pl. 27, figs. 15a–20c.
Globoturborotalita decoraperta (Takayanagi and Saito), 1962, p. 85, pl. 28, figs. 10a–10c.
Globoturborotalita druryi (Akers), 1955, p. 654, pl. 65, fig. 1.
Globoturborotalita nepenthes (Todd), 1957, p. 301, figs. 7a–7b.
Globoturborotalita obliquus (Bolli), 1957, p. 113, pl. 25, figs. 10a–10c.
Globoturborotalita woodi (Jenkins), 1960, p. 352, pl. 2, figs. 2a–2c.
Neogloboquadrina atlantica (Berggren), 1972, p. 979, pl. 2, fig. 8.
Neogloboquadrina acostaensis (Blow), 1959, p. 208, pl. 17, figs. 106a–106c.
Neogloboquadrina praeatlantica Foresi et al., 2002, p. 327, pl. 1, figs. 1–13; pl. 2, figs 1–12.
Neogloboquadrina pseudopachyderma Cita et al.), 1965, p. 233–236, pl. 20, figs. 3a–3c, 4a–4c, 6.
Orbulina suturalis Brönnimann, 1951, pt. 4, p. 135, Text fig. IV, figs. 15, 16, 20.
Orbulina universa d’Orbigny, 1839, p. 3, pl. 1, fig. 1.
Paragloborotalia continuosa (Blow), 1959, p. 218, pl. 19, figs. 125a–125c.
Paragloborotalia siakensis (LeRoy), 1939, p. 262, pl. 4, figs. 20–22.
Praeorbulina curva (Blow), 1956, p. 64, text fig. 1, no. 9–14.
Praeorbulina sicana (de Stefani), 1950, p. 9, fig. 6.
Sphaeroidinellopsis disjuncta Finlay, 1940, p. 467, pl. 67, figs. 224–228.
Sphaeroidinellopsis seminulina (Schwager), 1866, p. 256, pl. 7, fig. 112.
Sphaeroidinellopsis subdehiscens (Blow), 1959, p. 195, pl. 12, figs. 71a–72; Banner and Blow, 1960, p. 15, figs. 5a–5c.
Tenuitella clemenciae (Bermúdez), 1961, p.1321, pl. 17, fig. 10.
Tenuitella minuitissima (Bolli), 1957, p. 119, pl. 29, fig. 1.
1 Suzuki, T., Hayashi, H., and Idemitsu, K., 2020. Data report: middle Miocene planktonic foraminifers from the eastern equatorial Pacific, IODP Expedition 321 Site U1338. In Pälike, H., Lyle, M., Nishi, H., Raffi, I., Gamage, K., Klaus, A., and the Expedition 320/321 Scientists, Proceedings of the Integrated Ocean Drilling Program, 320/321: Tokyo (Integrated Ocean Drilling Program Management International, Inc.). https://doi.org/10.2204/iodp.proc.320321.221.2020
2 Interdisciplinary Faculty of Science and Engineering, Shimane University, Japan. Correspondence author: zaku142@gmail.com
This work is distributed under the Creative Commons Attribution 4.0 International (CC BY 4.0) license. 