Clemens, S.C., Kuhnt, W., LeVay, L.J., and the Expedition 353 Scientists
Proceedings of the International Ocean Discovery Program Volume 353
publications.iodp.org
doi:10.14379/iodp.proc.353.109.2016

Biostratigraphic summary1
M.M. Robinson, M. Bartol, C.T. Bolton, X. Ding, K. Gariboldi, O.E. Romero, and the Expedition 353 Scientists2
Keywords: International Ocean Discovery Program, IODP, Expedition 353, JOIDES Resolution, Site U1443, Site U1444, Site U1445, Site U1446, Site U1447, Site U1448, Indian monsoon, monsoon, Bay of Bengal, Andaman Sea, paleoclimate, paleoceanography, Miocene, Pliocene, Pleistocene, Holocene, Cretaceous, Paleogene, Ninetyeast Ridge, Indian Ocean, salinity, Bengal Fan, biostratigraphy, diatoms, planktonic foraminifers, calcareous nannofossils
MS 353-109: Published 29 July 2016
Introduction
A primary objective of International Ocean Discovery Program (IODP) Expedition 353 is to reconstruct changes in Indian monsoon circulation since the Miocene at tectonic to centennial timescales. During Expedition 353, six sites were drilled in the Bay of Bengal (Sites U1443–U1448), each strategically placed to capture the variable strength of the monsoon precipitation and runoff signature across the region. As a regional correlation among sites is integral to the reconstruction of monsoon changes through time, the shipboard micropaleontologists were called on to provide a biostratigraphically controlled age model for each site.
Although all investigated microfossil groups provided valuable information, calcareous nannofossils were critical to reducing the error surrounding species events, as multiple samples per core could be analyzed quickly. Planktonic foraminifers provided a suite of key datums at all sites, though error bars were sometimes large in zones of reworking in Site U1445 in the Mahanadi basin. The biostratigraphic utility of diatoms was at times hampered in the more calcareous sediments (Site U1443 on the Ninetyeast Ridge, Pliocene–Pleistocene of Site U1447, and Site U1448 posthiatus sequence) and during intervals of low productivity where abundances were low. The diatom datum events at Site U1445, however, were particularly useful.
Here, a synthesis of Expedition 353 biostratigraphy is presented. Future development of an oxygen isotope stratigraphy for these sites will provide an opportunity to improve the calibration of microfossil datum events.
Calcareous nannofossils
Overall, Miocene to Pleistocene calcareous nannofossil biostratigraphy in the Bay of Bengal appears to closely follow the global scheme in terms of event sequence (Gradstein et al., 2012). Sedimentation rates vary greatly among different sites (Figure F1), and the age of the oldest sediments reached at a particular site ranges between the Pleistocene and the Late Cretaceous. Sequences of nannofossil events recorded at different Expedition 353 sites thus cannot be directly compared; nevertheless, they appear to reflect a certain degree of regional variability. A few observations on calcareous nannofossils in the study region are noted below:
- Several Pleistocene bioevents are based on first occurrence, last occurrence, or first common occurrence of different size groups of Gephyrocapsa species. We found these events to be of limited use at all of the Expedition 353 sites. The reentrance of medium-sized Gephyrocapsa coccoliths (4–5.5 µm) was detected at several sites (often as the de facto first occurrence of the group) but was only distinct enough to be considered reliable at Site U1445. Gephyrocapsa coccoliths >5.5 µm were very rare and occurred mostly in the Upper Pleistocene above the horizon where their last occurrence was expected. Small Gephyrocapsa dominance was observed in many samples, yet the onset of this interval was not distinct at some sites and clearly diachronous among different sites.
- Reticulofenestra asanoi was only found at Andaman Sea Site U1447 and in one sample at Site U1448. This species was absent from studied samples at Ninetyeast Ridge and Mahanadi basin Sites U1443–U1446.
- In agreement with the findings of Flores et al. (2014), who did not find the middle Miocene marker species Helicosphaera ampliaperta in eastern Arabian Sea sediments, this species was absent from Bay of Bengal Site U1443. The species was detected at Andaman Sea Site U1448, where the top of its range is a clearly observable bioevent.
- Although usually present, the following marker species were always rare at Expedition 353 sites: Discoaster tamalis, Discoaster triradiatus, Discoaster asymmetricus, Amaurolithus spp., and Nicklithus amplificus.
- The deep photic zone–dwelling species Florisphaera profunda was very abundant in Pliocene–Pleistocene sediments at all Expedition 353 sites and especially at Andaman Sea Sites U1447 and U1448. The high degree of dominance of coccolith assemblages by F. profunda suggests that the water column remained strongly stratified throughout the Pliocene–Pleistocene, probably as a combined result of monsoon-related surface freshening and thermal stratification. These findings are consistent with a sediment trap study from the Bay of Bengal that suggests that the large component of deep production in this region drives higher organic carbon export compared to other stratified tropical ocean regions (Stoll et al., 2007). Future work will aim to quantify long-term changes in F. profunda abundance and provide a record of past stratification and productivity.
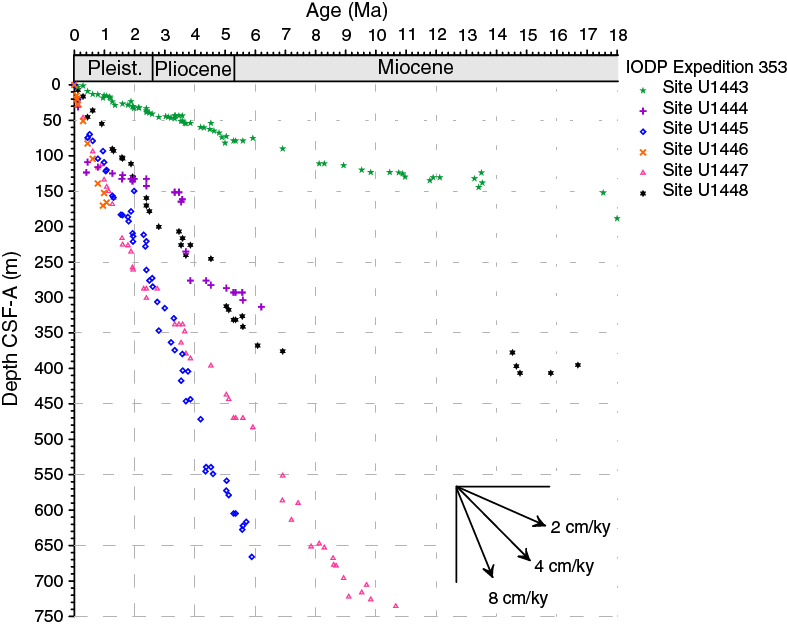
Figure F1. Neogene and Quaternary sedimentation rates for the six Expedition 353 sites in the Bay of Bengal.
Planktonic foraminifers
Several peculiarities regarding planktonic foraminifer species ranges and assemblage changes became apparent during this expedition. Together, they suggest unusual and evolving mixed layer environmental conditions characterizing the Bay of Bengal during the Pliocene:
- Although Globigerinoides ruber evolved in the late middle Miocene (~10 Ma; Kennett and Srinivasan, 1983) and is usually one of the most common species in warm subtropical to tropical assemblages, it does not appear in Expedition 353 sediments until the early Pliocene, after which it is consistently common. This delayed appearance is also noted in the Arabian Sea and is attributed to undetermined environmental conditions that do not affect any other species (Fleisher, 1974).
- The published first and last appearance datums of Globorotalia flexuosa are 0.40 and 0.07 Ma, respectively (Gradstein et al., 2012), making this species an important biostratigraphic marker in Pleistocene sediments. At Expedition 353 Sites U1443, U1445, U1447, and U1448, this species also appears in Pliocene sediments. G. flexuosa has also been documented in Pliocene sediments in the eastern tropical Indian Ocean (Shipboard Scientific Party, 1989a), in the western tropical Pacific (Kennett and Srinivasan, 1983), and in the Arabian Sea (Fleisher, 1974).
- The last appearance datum of Dentoglobigerina altispira is diachronous, occurring at different times in the Atlantic (3.13 Ma) and Pacific (3.47 Ma) Oceans (Gradstein et al., 2012). A younger age for this event is a better fit to our age models, particularly at Site U1445 where calcareous nannofossil, diatom, and magnetic events suggest an age near 3.00 Ma for this last occurrence, indicating an extended range of this species in the Bay of Bengal compared to the Atlantic and Pacific Oceans. In the Arabian Sea, rare appearances of D. altispira persist into the basal Pleistocene (Fleisher, 1974).
- Globigerina bulloides, Globigerinita glutinata, and Neogloboquadrina dutertrei are commonly associated, in the tropics, with cooler water and higher productivity that accompany upwelling zones. These species are consistently present in low to moderate percentages at the relatively shallow sites in the Andaman Sea and along the northeast Indian margin since the late Miocene. At open ocean Sites U1443 and U1444, however, these species do not appear until ~3 Ma, at which time they become a sustained and important part of the assemblage. Concurrently, their relative abundance increases at Site U1445. This change may indicate the onset of vertical mixing of intermediate and deep waters in the central Bay of Bengal that began ~3 Ma.
Together, the delayed occurrence of G. ruber until the early Pliocene, the early appearance of G. flexuosa in this region during the Pliocene, the extended range of D. altispira, and the inclusion of G. bulloides, G. glutinata, and N. dutertrei in the central Bay of Bengal at ~3 Ma describe a unique and evolving sea-surface environment leading into and during the Pliocene. We suspect these changes reflect evolving monsoon behavior during this time, though continued onshore research will be required to document the connection.
Diatoms
Diatom bioevents in the Eastern Indian Ocean mostly agree with those documented in the Pacific Ocean and equatorial Indian Ocean (Barron, 1985a, 1985b; Mikkelsen, 1990; Fourtanier, 1991). However, the occurrence of some index species, such as Rocella spp., will be further investigated to clarify whether they have a different range in the Bay of Bengal. Sporadic diatom bioevents were useful for stratigraphy. This limited utility is mainly due to their discontinuous occurrence and the rare presence of index species in the Bay of Bengal Cenozoic sediments.
The study of diatoms in Expedition 353 downcore sediments will contribute to the comprehension of the paleoceanography and paleoproductivity changes that occurred through the Neogene in the Bay of Bengal. Whenever present in abundance, the diatom community is highly diverse, consisting mainly of species typical of warm to temperate low-latitude ocean waters. High-productivity species, including Thalassionema nitzschioides var. nitzschioides and resting spores of Chaetoceros, tend to dominate whenever total diatom abundance is higher than “few.” Valve preservation ranges from good to poor and tends to be better whenever abundance is higher.
On the regional scale, three diatom-rich time intervals were identified:
- Pleistocene: diatoms are present at all the investigated sites (except Site U1444).
- Pliocene: diatoms dominate at Site U1445; several bloom-like, diatom-dominated events were recorded.
- Late Miocene: diatoms are present at Sites U1445 (Mahanadi basin) and U1447 and in the prehiatus sequence of Site U1448 (Andaman Sea).
Diatoms are more abundant at coastal sites (Mahanadi basin Sites U1445–U1447 and Andaman Sea Site U1448) than at pelagic Sites U1443 and U1444. Site U1445 is the most diatom-rich site, as diatoms are present throughout the entire recovered sedimentary record. At this site, the presence of several freshwater taxa (to include Aulacoseira, Epithemia, Navicula, Pinnularia, and Stephanodiscus) and a diverse phytolith community reveal a certain degree of freshwater input through winds or river runoff. Sites U1443 and U1444, the farthest from land, are the most diatom poor.
Our observations at Site U1443 partially match those generated earlier at Ocean Drilling Program Site 758 (Ninetyeast Ridge). Diatoms, although often rare, occurred from the Pleistocene to the Campanian at Site 758 (Shipboard Scientific Party, 1989b; Fourtanier, 1991). We found, however, a less continuous record of diatom presence than previously reported at Site 758; diatoms at Site U1443 were found only in the Pleistocene and sporadically in the middle and early Miocene.
Sedimentation rates
The biochronologic analysis of the six Expedition 353 sites indicates an overall remarkably linear sedimentation rate pattern in the Bay of Bengal, occasionally interrupted by both small (Sites U1443 and U1447) and large (Sites U1443 and U1448) hiatuses as well as small and large turbiditic intervals at Sites U1447 and U1444, respectively. Biochronologic age estimates and sedimentation rate patterns are in close agreement with the magnetostratigraphy developed on board the ship. Neogene and Quaternary sedimentation rates for the six Expedition 353 sites are compared in Figure F1.
References
Barron, J.A., 1985a. Late Eocene to Holocene diatom biostratigraphy of the equatorial Pacific Ocean, Deep Sea Drilling Project Leg 85. In Mayer, L., Theyer, F., Thomas, E., et al., Initial Reports of the Deep Sea Drilling Project, 85: Washington, DC (U.S. Government Printing Office), 413–456. http://dx.doi.org/10.2973/dsdp.proc.85.108.1985
Barron, J.A., 1985b. Miocene to Holocene planktic diatoms. In Bolli, H.M., Saunders, J.B., and Perch-Nielsen, K. (Eds.), Plankton Stratigraphy: Cambridge, United Kingdom (Cambridge University Press), 763–809.
Fleisher, R.L., 1974. Cenozoic planktonic foraminifera and biostratigraphy, Arabian Sea Deep Sea Drilling Project, Leg 23A. In Whitmarsh, R.B., Weser, O.E., Ross, D.A., et al., Initial Reports of the Deep Sea Drilling Project, 23: Washington, DC (U.S. Govt. Printing Office), 1001–1072. http://dx.doi.org/10.2973/dsdp.proc.23.139.1974
Flores, J.A., Johnson, J.E., Mejía-Molina, A.E., Álverez, M.C., Sierro, F.J., Singh, S.D., Mahanti, S., and Giosan, L., 2014. Sedimentation rates from calcareous nannofossil and planktonic foraminifera biostratigraphy in the Andaman Sea, northern Bay of Bengal, and eastern Arabian Sea. Marine and Petroleum Geology, 58(Part A):425–437. http://dx.doi.org/10.1016/j.marpetgeo.2014.08.011
Fourtanier, E., 1991. Diatom biostratigraphy of equatorial Indian Ocean Site 758. In Weissel, J., Peirce, J., Taylor, E., Alt, J., et al., Proceedings of the Ocean Drilling Program, Scientific Results, 121: College Station, TX (Ocean Drilling Program), 189–208. http://dx.doi.org/10.2973/odp.proc.sr.121.137.1991
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. (Eds.), 2012. The Geological Time Scale 2012: Amsterdam (Elsevier).
Kennett, J.P., and Srinivasan, M.S., 1983. Neogene Planktonic Foraminifera: A Phylogenetic Atlas: Stroudsburg, PA (Hutchinson Ross).
Mikkelsen, N., 1990. Cenozoic diatom biostratigraphy and paleoceanography of the western equatorial Indian Ocean. In Duncan, R.A., Backman, J., Peterson, L.C., et al., Proceedings of the Ocean Drilling Program, Scientific Results, 115: College Station, TX (Ocean Drilling Program), 411–432. http://dx.doi.org/10.2973/odp.proc.sr.115.157.1990
Shipboard Scientific Party, 1989a. Site 757. In Peirce, J., Weissel, J., et al., Proceedings of the Ocean Drilling Program, Initial Reports, 121: College Station, TX (Ocean Drilling Program), 305–358. http://dx.doi.org/:10.2973/odp.proc.ir.121.111.1989
Shipboard Scientific Party, 1989b. Site 758. In Peirce, J., Weissel, J., et al., Proceedings of the Ocean Drilling Program, Initial Reports, 121: College Station, TX (Ocean Drilling Program), 359–453. http://dx.doi.org/10.2973/odp.proc.ir.121.112.1989
Stoll, H.M., Arevalos, A., Burke, A., Ziveri, P., Mortyn, G., Shimizu, N., and Unger, D., 2007. Seasonal cycles in biogenic production and export in northern Bay of Bengal sediment traps. Deep-Sea Research, Part II, 54(5–7):558–580. http://dx.doi.org/10.1016/j.dsr2.2007.01.002
1Robinson, M.M., Bartol, M., Bolton, C.T., Ding, X., Gariboldi, K., Romero, O.E., and the Expedition 353 Scientists, 2016. Biostratigraphic summary. In Clemens, S.C., Kuhnt, W., LeVay, L.J., and the Expedition 353 Scientists, Indian Monsoon Rainfall. Proceedings of the International Ocean Discovery Program, 353: College Station, TX (International Ocean Discovery Program). http://dx.doi.org/10.14379/iodp.proc.353.109.2016