Morgan, J., Gulick, S., Mellett, C.L., Green, S.L., and the Expedition 364 Scientists
Proceedings of the International Ocean Discovery Program Volume 364
publications.iodp.org
https://doi.org/10.14379/iodp.proc.364.108.2017

Site M0077: microbiology1
S. Gulick, J. Morgan, C.L. Mellett, S.L. Green, T. Bralower, E. Chenot, G. Christeson, P. Claeys, C. Cockell, M.J.L. Coolen, L. Ferrière, C. Gebhardt, K. Goto, H. Jones, D. Kring, J. Lofi, C. Lowery, R. Ocampo-Torres, L. Perez-Cruz, A.E. Pickersgill, M. Poelchau, A. Rae, C. Rasmussen, M. Rebolledo-Vieyra, U. Riller, H. Sato, J. Smit, S. Tikoo, N. Tomioka, J. Urrutia-Fucugauchi, M. Whalen, A. Wittmann, K. Yamaguchi, L. Xiao, and W. Zylberman2
Keywords: International Ocean Discovery Program, IODP, International Continental Scientific Drilling Program, ICDP, L/B Myrtle, Mission Specific Platform, Expedition 364, Site M0077, Hole M0077A, Gulf of México, Yucatán shelf, Chicxulub, impact crater, crater modification, multi-ring basin, peak ring, uplifted continental crust, impact melt rock, planar deformation features, Cretaceous/Paleogene boundary, PETM, K-Pg boundary, Cretaceous-Paleogene mass extinction, shock metamorphism, carbon isotope excursions, hydrothermal, geomagnetic reversal, shatter cone, ejecta, suevite, granitoid, pelagic limestone, tsunamite
MS 364-108: Published 30 December 2017
Geomicrobiology objectives
Asteroid and comet impacts are known to cause severe disruption to surface biota, as is the case for Chicxulub, but what is their effect on the present and ancient deep biosphere, where a large proportion of the Earth’s biomass resides?
Present biosphere
One of the objectives of sampling the core during Expedition 364 was to determine the present-day abundance and diversity of the subsurface biosphere that allows comparisons to the relatively well characterized deep marine sediment biosphere (Kallmeyer et al., 2012). Specific objectives were to (1) determine biomass with depth, (2) attempt cultivation of microbial taxa through the core, and (3) extract DNA to study the identity and diversity of the microbial community throughout the core.
Impacts can fracture rocks, increasing their porosity and permeability. For example, it has been demonstrated that the effects of high shock pressures on gneissic rocks in the 24 km diameter Haughton crater in the Canadian Arctic have made them more porous and translucent and therefore more colonizable by cyanobacteria (Cockell et al., 2002). Drilling of the Chesapeake Bay impact crater has shown a deep region of impact-fractured rocks (Cockell et al., 2012) where microbial abundance considerably increases in a region of the core that exhibits evidence for elevated temperatures. This finding suggests that once the impact-induced hydrothermal system cooled, conditions improved to support deep subsurface recolonization.
The brecciated regions within impact craters are of particular interest. Heavily fractured, brecciated rock is likely to improve fluid flow and the availability of nutrients and redox couples to microbiota. The geochemical diversity of different rocks shattered and mixed within the breccia layer will also increase the potential for diverse geochemical gradients, which are known to be beneficial for microorganisms in the deep biosphere (Roussel et al., 2008).
Finally, the geomicrobiology results at Chicxulub have astrobiological significance, such as understanding the effect of impacts on habitats for early life on Earth, when the impact flux was much higher than today, and the effect of impacts on the habitability of the Martian deep subsurface.
Ancient biosphere
Another objective is to study the ancient deep biosphere within the recovered core. The current state of knowledge is that ancient planktic DNA can be recovered from sediments as old as 270,000 y and successfully used for ecosystem reconstructions (Coolen and Overmann, 2007; Randlett et al., 2014). The majority of DNA in the marine sedimentary record was reported to be extracellular (Corinaldesi et al., 2008) and protected against microbial enzymatic attack through mineral adsorption. Mineral adsorption also prevents vertical migration, so this pool of immobilized DNA can be used for stratigraphic analysis (Coolen et al., 2006). Low-energy availability and subsequent ultraslow growth of microbial communities in the deep biosphere could further aid the preservation of DNA through deep time. However, it remains unknown to what extent ancient plankton DNA can be recovered from the deep biosphere and whether it is of use for deep-time ecosystem reconstructions, notably across the major extinction events. For this project, we will explore to what extent ancient planktic DNA can be recovered from the post-impact sedimentary rocks overlying the Chicxulub impact crater and whether this DNA is suitable for study of post-impact recovery and development of marine life.
In addition, we will explore to what extent bacteria and/or archaea in the post-impact sedimentary rock record can provide information about post-impact environmental and depositional conditions (e.g., sea-surface temperature and salinity, as well as sea level changes), based on the assumption that a substantial part of the microbiota in the deep biosphere was originally seeded from the ancient water column above (Kallmeyer et al., 2012). Subsurface microbes that became energy limited upon burial but remained capable of staying alive (e.g., through DNA repair; Hoehler and Jorgensen, 2013) must have evolved slowly due to growth limitations (Orsi et al., 2013) and are hence expected to show a high degree of taxonomic similarity to microbes found in similar environments today. For example, this hypothesis may explain why the microbial diversity in a 20 My old coal deposit buried underneath 2.5 km of marine sediments still resembled microbial taxa typically found in modern forest soils (Inagaki et al., 2015).
Depositional paleoenvironment associated with the ancient biosphere
Another goal is to study biomarkers to investigate life and environmental conditions within the post-impact ocean. Biomarker or molecular fossils are organic compounds of biosynthetic origin and can be preserved in geological records for hundreds of millions of years. The composition of the fossil biomarkers can reveal information about ancient biodiversity and complement stable isotopic data in establishing the paleoenvironmental conditions that prevailed at the time of deposition such as trophic status, euxinia, ocean acidification, changes in hydrological balance, and atmospheric CO2 concentrations (Brocks and Grice, 2011; Brocks and Summons, 2014; Whiteside and Grice, 2016). Lipid biomarkers originate from all three domains of microbial life: eukaryotes, bacteria, and archaea (Brocks and Grice, 2011; Brocks and Summons, 2014; Grice and Brocks, 2011; Grice and Eiserbeck, 2014; Peters et al., 2004; Whiteside and Grice, 2016). Their chemical structure can be related directly or indirectly to biological precursors and can be specific or nonspecific in terms of source organisms (Brocks and Grice, 2011). For example, the paired analysis of more recalcitrant lipid biomarkers could reveal whether life was present in the past if the more labile nucleic acids and/or microbial cells are below the detection limit.
In summary, the following research questions will be addressed in postexpedition research concerning the ancient biosphere and associated paleoenvironmental conditions:
- What is the developmental history of planktic taxa and associated biomarkers in the post-impact ocean basin?
- To what extent does the taxonomic and functional diversity of possibly slow-evolving subsurface bacteria correlate with changes in paleoenvironmental conditions or assist in the reconstruction of the paleodepositional environment?
Drilling fluid contaminants
The aforementioned microbiology goals for Expedition 364 will be carried out postexpedition. Reported here are the cell counts completed within the drilling fluid samples that were collected alongside samples used for scientific analysis (Smith et al., 2000a, 2000b). The purpose was to allow for the enumeration and anaerobic cultivation of contaminant microbes in the drilling fluid and the extraction of nucleic acids from drilling fluid to compare to interior nucleic acid samples. Figure F1 shows the cell count in the drilling fluid throughout the cored interval of Expedition 364, which can be compared with cell counts within the microbiology samples.
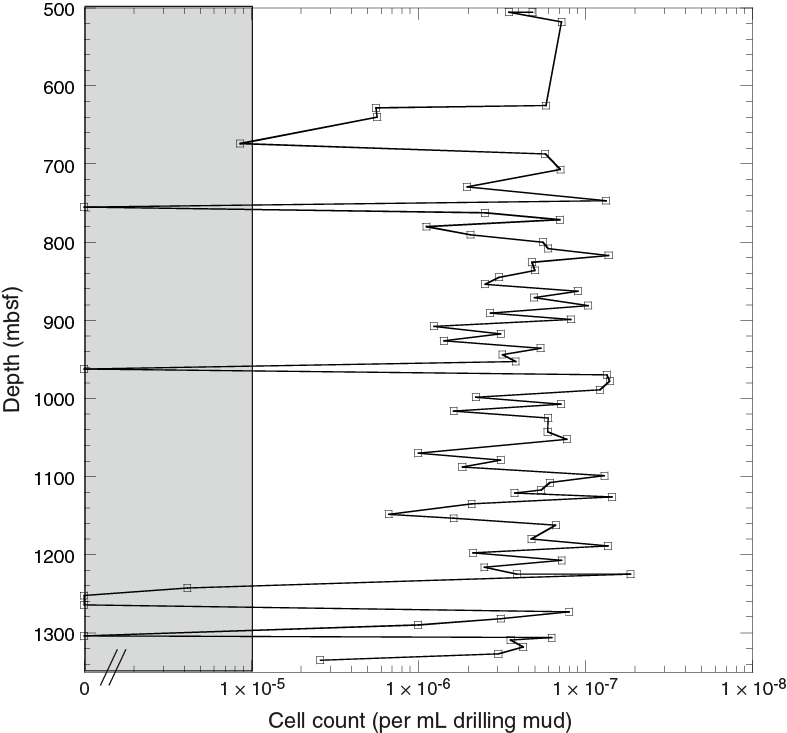
Figure F1. Microbial enumerations determined in drilling fluid.
References
Brocks, J.J., and Grice, K., 2011. Biomarkers (molecular fossils). In Reitner, J., and Thiel, V. (Eds.), Encyclopedia of Geobiology: Dordrecht, The Netherlands (Springer), 147–167. https://doi.org/10.1007/978-1-4020-9212-1_30
Brocks, J.J., and Summons, R.E., 2014. Sedimentary hydrocarbons, biomarkers for early life. In Karl, D.M., and Schlesinger, W.H. (Eds.), Treatise on Geochemistry (2nd edition) (Volume 10): Biogeochemistry. Holland, H., and Turekian, K.K. (Series Eds.): Oxford, United Kingdom (Elsevier), 61–103. https://doi.org/10.1016/B978-0-08-095975-7.00803-2
Cockell, C.S., Lee, P., Osinski, G., Horneck, G., and Broady, P., 2002. Impact-induced microbial endolithic habitats. Meteoritics & Planetary Science, 37(10):1287–1298. https://doi.org/10.1111/j.1945-5100.2002.tb01029.x
Cockell, C.S., Voytek, M.A., Gronstal, A.L., Finster, K., Kirshtein, J.D., Howard, K., Reitner, J., Gohn, G.S., Sanford, W.E., Horton, J.W., Jr., Kallmeyer, J., Kelly, L., and Powars, D.S., 2012. Impact disruption and recovery of the deep subsurface biosphere. Astrobiology, 12(3):231–246. https://doi.org/10.1089/ast.2011.0722
Coolen, M.J.L., Boere, A., Abbas, B., Baas, M., Wakeham, S.G., and Sinninghe Damsté, J.S., 2006. Ancient DNA derived from alkenone-biosynthesizing haptophytes and other algae in Holocene sediments from the Black Sea. Paleoceanography, 21(1):PA1005. https://doi.org/10.1029/2005PA001188
Coolen, M.J.L., and Orsi, W.D., 2015. The transcriptional response of microbial communities in thawing Alaskan permafrost soils. Frontiers in Microbiology, 6:197. https://doi.org/10.3389/fmicb.2015.00197
Coolen, M.J.L., and Overmann, J., 2007. 217 000-year-old DNA sequences of green sulfur bacteria in Mediterranean sapropels and their implications for the reconstruction of the paleoenvironment. Environmental Microbiology, 9(1):238–249. https://doi.org/10.1111/j.1462-2920.2006.01134.x
Corinaldesi, C., Beolchini, F., and Dell’Anno, A., 2008. Damage and degradation rates of extracellular DNA in marine sediments: implications for the preservation of gene sequences. Molecular Ecology, 17(17):3939–3951. https://doi.org/10.1111/j.1365-294X.2008.03880.x
Grice, K., and Brocks, J.J., 2011. Biomarkers (organic, compound-specific isotopes). In Reitner, J., and Thiel, V. (Eds.), Encyclopedia of Geobiology: Dordrecht, The Netherlands (Springer), 167–182. https://doi.org/10.1007/978-1-4020-9212-1_29
Grice, K., and Eiserbeck, C., 2014. The analysis and application of biomarkers. In Falkowski, P.G., and Freeman, K.H. (Eds.), Treatise on Geochemistry (2nd edition) (Volume 12): Organic Geochemistry. Holland, H., and Turekian, K.K. (Series Eds.): Oxford, United Kingdom (Elsevier), 47–78. https://doi.org/10.1016/B978-0-08-095975-7.01006-8
Gulick, S., Morgan, J., Mellett, C.L., Green, S.L., Bralower, T., Chenot, E., Christeson, G., Claeys, P., Cockell, C., Coolen, M.J.L., Ferrière, L., Gebhardt, C., Goto, K., Jones, H., Kring, D., Lofi, J., Lowery, C., Ocampo-Torres, R., Perez-Cruz, L., Pickersgill, A.E., Poelchau, M., Rae, A., Rasmussen, C., Rebdolledo-Vieyra, M., Riller, U., Sato, H., Smit, J., Tikoo, S., Tomioka, N., Urrutia Fucugauchi, J., Whalen, M., Wittmann, A., Yamagachi, K., Xiao, L., and Zylberman, W., 2017. Expedition 364 methods. In Morgan, J., Gulick, S., Mellett, C.L., Green, S.L., and the Expedition 364 Scientists, Chicxulub: Drilling the K-Pg Impact Crater. Proceedings of the International Ocean Discovery Program, 364: College Station, TX (International Ocean Discovery Program). https://doi.org/10.14379/iodp.proc.364.102.2017
Hoehler, T.M., and Jørgensen, B.B., 2013. Microbial life under extreme energy limitation. National Review of Microbiology, 11(2):83–94. https://doi.org/10.1038/nrmicro2939
Inagaki, F., Hinrichs, K.-U., Kubo, Y., Bowles, M.W., Heuer, V.B., Long, W.-L., Hoshino, T., Ijiri, A., Imachi, H., Ito, M., Kaneko, M., Lever, M.A., Lin, Y.-S., Methé, B.A., Morita, S., Morono, Y., Tanikawa, W., Bihan, M., Bowden, S.A., Elvert, M., Glombitza, C., Gross, D., Harrington, G.J., Hori, T., Li, K., Limmer, D., Liu, C.-H., Murayama, M., Ohkouchi, N., Ono, S., Park, Y.-S., Phillips, S.C., Prieto-Mollar, X., Purkey, M., Riedinger, N., Sanada, Y., Sauvage, J., Snyder, G., Susilawati, R., Takano, Y., Tasumi, E., Terada, T., Tomaru, H., Trembath-Reichert, E., Wang, D.T., and Yamada, Y., 2015. Exploring deep microbial life in coal-bearing sediment down to ~2.5 km below the ocean floor. Science, 349(6246):420–424. http://dx.doi.org/10.1126/science.aaa6882
Kallmeyer, J., Pockalny, R., Adhikari, R.R., Smith, D.C., and D’Hondt, S., 2012. Global distribution of microbial abundance and biomass in subseafloor sediment. Proceedings of the National Academy of Sciences of the United States of America, 109(40):16213–16216. http://dx.doi.org/10.1073/pnas.1203849109
Orsi, W.D., Edgcomb, V.P., Christman, G.D., and Biddle, J.F., 2013. Gene expression in the deep biosphere. Nature, 499(7457):205–208. http://dx.doi.org/10.1038/nature12230
Peters, K.E., Walters, C.C., and Moldowan, J.M., 2004. The Biomarker Guide (Volume 2): Biomarkers and Isotopes in Petroleum Systems and Earth History: Cambridge, United Kingdom (Cambridge University Press). https://doi.org/10.1017/CBO9781107326040
Randlett, M.-È., Coolen, M.J.L., Stockhecke, M., Pickarski, N., Litt, T., Balkema, C., Kwiecien, O., Tomonaga, Y., Wehrli, B., and Schubert, C.J., 2014. Alkenone distribution in Lake Van sediment over the last 270 ka: influence of temperature and haptophyte species composition. Quaternary Science Reviews, 104:53–62. https://doi.org/10.1016/j.quascirev.2014.07.009
Roussel, E.G., Cambon Bonavita, M.-A., Querellou, J., Cragg, B.A., Webster, G., Prieur, D., and Parkes, J.R., 2008. Extending the sub-sea-floor biosphere. Science, 320(5879):1046. http://dx.doi.org/10.1126/science.1154545
Smith, D.C., Spivack, A.J., Fisk, M.R., Haveman, S.A., and Staudigel, H., 2000a. Tracer-based estimates of drilling-induced microbial contamination of deep sea crust. Geomicrobiology Journal, 17(3):207–219. http://dx.doi.org/10.1080/01490450050121170
Smith, D.C., Spivack, A.J., Fisk, M.R., Haveman, S.A., Staudigel, H., and the Leg 185 Shipboard Scientific Party, 2000b. Technical Note 28: Methods for Quantifying Potential Microbial Contamination during Deep Ocean Coring. Ocean Drilling Program. http://dx.doi.org/10.2973/odp.tn.28.2000
Whiteside, J.H., and Grice, K., 2016. Biomarker records associated with mass extinction events. Annual Review of Earth and Planetary Sciences, 44(1):581–612. https://doi.org/10.1146/annurev-earth-060115-012501
1 Gulick, S., Morgan, J., Mellett, C.L., Green, S.L., Bralower, T., Chenot, E., Christeson, G., Claeys, P., Cockell, C., Coolen, M.J.L., Ferrière, L., Gebhardt, C., Goto, K., Jones, H., Kring, D., Lofi, J., Lowery, C., Ocampo-Torres, R., Perez-Cruz, L., Pickersgill, A.E., Poelchau, M., Rae, A., Rasmussen, C., Rebolledo-Vieyra, M., Riller, U., Sato, H., Smit, J., Tikoo, S., Tomioka, N., Urrutia- Fucugauchi, J., Whalen, M., Wittmann, A., Yamaguchi, K., Xiao, L., and Zylberman, W., 2017. Site M0077: microbiology. In Morgan, J., Gulick, S., Mellett, C.L., Green, S.L., and the Expedition 364 Scientists, Chicxulub: Drilling the K-Pg Impact Crater. Proceedings of the International Ocean Discovery Program, 364: College Station, TX (International Ocean Discovery Program). https://doi.org/10.14379/iodp.proc.364.108.2017
2 Expedition 364 Scientists’ addresses.
This work is distributed under the Creative Commons Attribution 4.0 International (CC BY 4.0) license. 