Heuer, V.B., Inagaki, F., Morono, Y., Kubo, Y., Maeda, L., and the Expedition 370 Scientists
Proceedings of the International Ocean Discovery Program Volume 370
publications.iodp.org
https://doi.org/10.14379/iodp.proc.370.201.2018

Data report: calcareous nannofossils from the middle Miocene to Pleistocene, IODP Expedition 370 Site C00231
Kyoko Hagino2 and the Expedition 370 Scientists3
Keywords: International Ocean Discovery Program, IODP, Chikyu, Expedition 370, Site C0023, calcareous nannofossil, biostratigraphy, Neogene
MS 370-201: Received 26 August 2017 · Accepted 13 December 2017 · Published 18 April 2018
Abstract
International Ocean Discovery Program Expedition 370 recovered marine subsurface sediments from 189 to 1126 meters below seafloor (mbsf) at Site C0023, which was established on the Nankai Trough, off Muroto Peninsula, Japan. Biostratigraphic studies based on calcareous nannofossil assemblages revealed the ages of sediments are from the middle Miocene to Pleistocene. Samples from 1126.885 to 796.530 mbsf yielded Miocene taxa such as Coccolithus miopelagicus, Cyclicargolithus floridanus, Catinaster calyculus, and Discoaster quinqueramus. The interval 790.285–774.984 mbsf spans the late Miocene to the early Pliocene, as it corresponds to bioevents spanning from the last occurrence (LO) of D. quinqueramus to the first occurrence of Ceratolithus cristatus. Samples from 769.440 to 639.982 mbsf are referred to the Pliocene based on occurrences of C. cristatus, Reticulofenestra pseudoumbilicus (≥7 µm), and/or Discoaster surculus. Sediments from 630.440 to 190.483 mbsf are dated to the Pleistocene, as an interval above the LO of Discoaster pentaradiatus. Preservation of calcareous nannofossils is poor or moderate in the middle Miocene samples and moderate to good in the samples from the upper Miocene to Pleistocene.
Introduction
During International Ocean Discovery Program (IODP) Expedition 370, Site C0023 was drilled at 32°22.00′N, 134°57.98′E in 4776 m water depth in the northwest Pacific Ocean off Kochi, Japan (Figure F1). Site C0023 is located on the protothrust zone of the Nankai accretionary complex, in the vicinity of Ocean Drilling Program (ODP) Sites 808 and 1174. The Nankai accretionary complex comprises a thick sediment pile consisting of turbidite layers overlaying hemipelagic sediment accumulated on the young (~16 Ma) basaltic basement (e.g., Shipboard Scientific Party, 1991, 2001). The sedimentation rate based on biostratigraphy for the late Quaternary Nankai Trough deposit was significantly high: 800–1350 m/My in the last 0.46 My at Site 808 and 630–770 m/My in the last 0.08 My at the Site 1174 (Olafsson, 1993; Shipboard Scientific Party, 2001). On the other hand, the sedimentation rate for the hemipelagic Shikoku Basin deposits was relatively low: 24–200 m/My for sediments older than 0.46 Ma at Site 808 and 11–125 m/My for sediments older than 0.08 Ma at Site 1174 (Olafsson, 1993; Shipboard Scientific Party, 2001). The lowermost sediments above the basaltic basement were referred to middle Miocene calcareous nannofossil Zone NN5 at Site 808 (Shipboard Scientific Party, 2001).
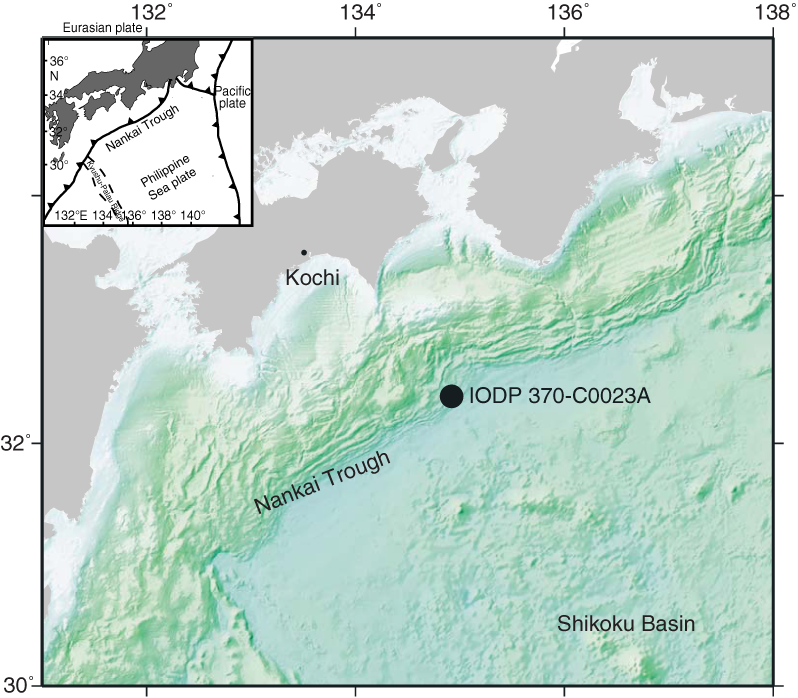
Figure F1. Location of Hole C0023A.
The main objectives of this expedition were to (1) explore the limit of distribution of microbial communities in marine subsurface sediments, (2) reveal geochemical, geophysical, and hydrogeological characteristics of the subsurface sediments, and (3) examine underlying basaltic basement of the Nankai accretionary complex (see the Expedition 370 summary chapter [Heuer et al., 2017]). Calcareous nannofossil assemblages in the recovered sediments were studied in order to determine the ages of sediments. This data report documents the middle Miocene to Pleistocene calcareous nannofossil assemblages and assigned biostratigraphic zones recorded for this site.
Methods and materials
Hole C0023A was washed down to 181 meters below seafloor (mbsf) and then cored from 189 to 1180 mbsf using the hydraulic piston coring, extended shoe coring, and rotary core barrel systems (Figure F1) (see the Expedition 370 summary chapter [Heuer et al., 2017]). Smear slides were prepared for the study of calcareous nannofossils from sediment samples obtained from core catcher samples or from the bottoms of cores using standard techniques (e.g., Bown and Young, 1998). Calcareous nannofossils were examined at 1500× magnification under a Nikon E600 polarizing light microscope.
Nannofossil preservation was recorded as follows:
- G = good (little or no evidence of dissolution and/or overgrowth, little or no alteration of primary morphological features, and specimens are identifiable to the species level).
- M = moderate (minor dissolution or crystal overgrowth observed, some alteration of primary morphological features, but most specimens are identifiable to the species level).
- P = poor (strong dissolution or crystal overgrowth, significant alteration of primary morphological features, and many specimens are unidentifiable at the species and/or generic level).
The abundance of total calcareous nannofossils and individual species for each sample was estimated as follows:
- A = abundant (11 or more specimens per field of view).
- C = common (1–10 specimens per field of view).
- F = few (1 specimen per 2–10 fields of view).
- R = rare (1 specimen per 11–30 fields of view).
- VR = very rare (1 specimen per 31 or more fields of view).
- B = barren.
Results were correlated to the calcareous nannofossil biostratigraphic NN zones of Martini (1971) and CN zones of Okada and Bukry (1980). Furthermore, absolute ages for datums and additional biostratigraphic events were assigned on the basis of Backman et al. (2012) whenever possible. Gephyrocapsa spp. and Reticulofenestra spp. were classified to groups on the basis of coccolith size following Backman et al. (2012) and Young (1998). Calcareous nannofossil species considered in this study are listed in the Appendix. Bibliographic references for these taxa can be found in Perch-Nielsen (1985) and Bown (1998). In order to determine the onset of the acme of Emiliania huxleyi, 300 specimens of calcareous nannofossils other than Florisphaera profunda were identified in the samples that contained E. huxleyi. Age of onset of the acme of E. huxleyi followed Raffi et al. (2006).
Results
Biostratigraphy
A total of 88 out of 112 studied samples yielded sufficient numbers of calcareous nannofossils for biostratigraphic studies. Ages of sediments range from middle Miocene to early Pleistocene. Preservation of calcareous nannofossils is poor or moderate in middle Miocene samples and moderate to good in samples from the upper Miocene to Pleistocene (Table T1). The bottom two samples (370-C0023A-112R-1, 48.0–50.0 cm, and 111R-CC, 46.0–48.0 cm [1177.000–1173.980 mbsf]) are basaltic basement (see the Expedition 370 summary chapter [Heuer et al., 2017]) and did not yield any calcareous nannofossils.
Table T1. Calcareous nannofossil occurrences. Download table in CSV format.
Interval 110R-CC, 32.0–34.0 cm, through 88R-CC, 47.5–49.5 cm (1126.885–914.565 mbsf), yielded a middle Miocene calcareous nannofossil assemblage characterized by Coccolithus miopelagicus and Cyclicargolithus floridanus (Plate P1, figures 1, 2). In this interval, samples from interval 110R-CC, 32.0–34.0 cm, through 105R-CC, 33.0–35.0 cm (1126.885–1084.100 mbsf), were assigned to Zone NN5 (CN4) on the basis of the occurrence of Sphenolithus heteromorphus (17.75–13.53 Ma; Plate P1, figures 3a, 3b). Zone NN6 (CN5a) is a partial range zone that extends from the last occurrence (LO) of S. heteromorphus (13.53 Ma) to the first occurrence (FO) of Discoaster kugleri (11.88 Ma) (Young, 1998; Backman et al., 2012). Because D. kugleri was not recognized at this site, the top of Zone NN6 (CN5a) was approximated based on the LO of C. floridanus (~12.0 Ma) between Samples 94R-CC, 33.5–35.5 cm, and 91R-CC, 43.0–45.0 cm (977.981–947.000 mbsf). Zone NN7 (CN5b) is a successive first appearance zone defined by the FO of D. kugleri (base of Zone NN7) and the FO of Catinaster coalitus (10.79 Ma) (Young, 1998; Backman et al., 2012). This site yielded neither D. kugleri nor C. coalitus; therefore, Zone NN7 (CN5b) was assigned to between Samples 91R-CC, 43.0–45.0 cm, and 88R-CC, 47.5–49.5 cm (947.00–914.565 mbsf), based on the LO of C. floridanus (~12.0 Ma) and the LO of C. miopelagicus (~10.61 Ma).
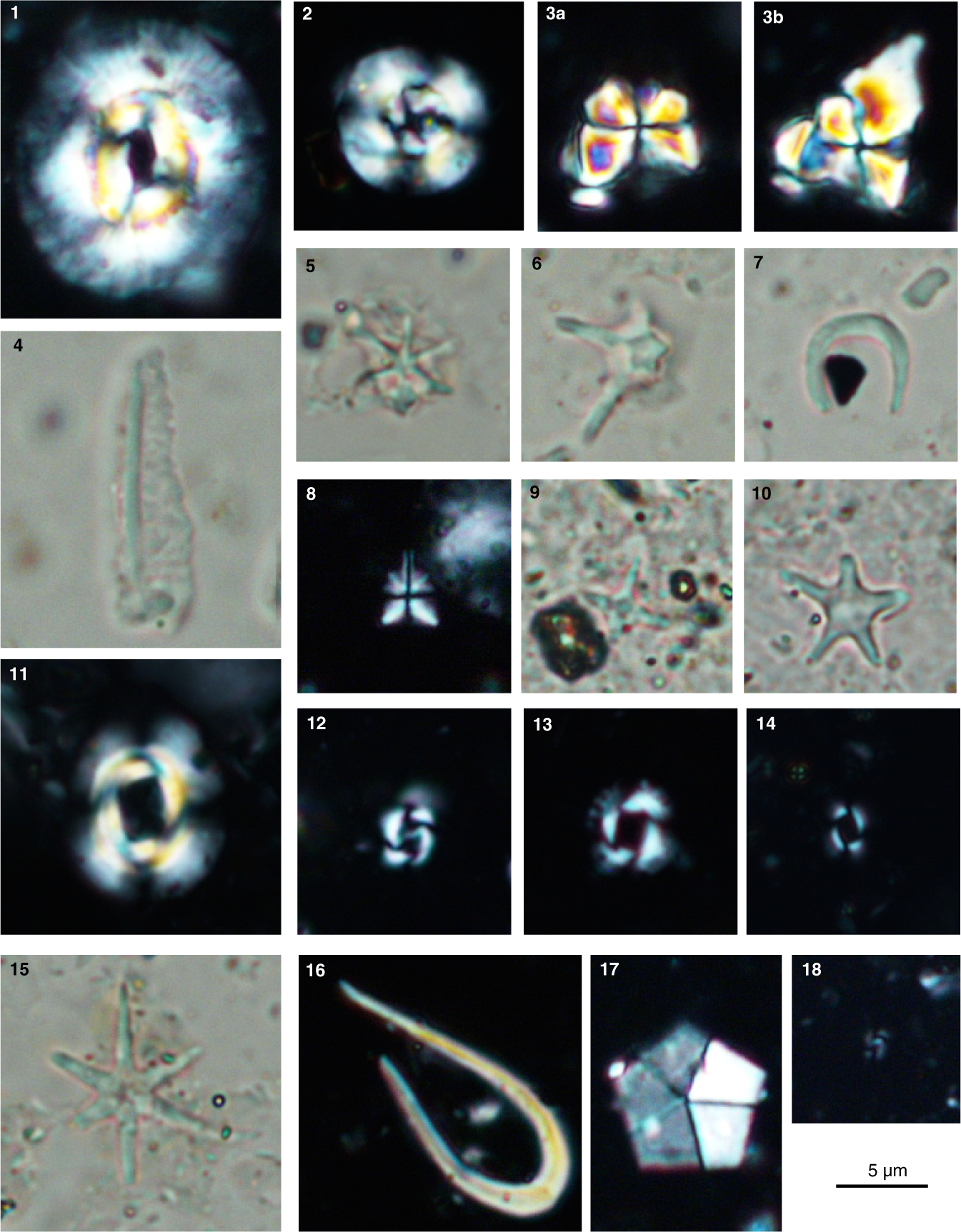
Plate P1. Light microscopic images of calcareous nannofossils.
Sample 82R-CC, 28.5–30.5 cm (856.175–856.195 mbsf), is assigned to late Miocene Zones NN9–NN10a (CN7b–CN8a) based on the occurrence of Catinaster calyculus (10.71–9.65 Ma; Plate P1, figure 5). The interval between Samples 79R-CC, 13.0–15.0 cm, and 71R-CC, 13.5–15.5 cm (832.525–796.530 mbsf), was defined as late Miocene Zone NN11 (CN9) on the basis of the FO of Discoaster berggrenii (8.2 Ma; Plate P1, figure 6) and the LO of Discoaster quinqueramus (5.53 Ma).
Zone NN12 (CN10a–CN10b) spans the late Miocene to early Pliocene and was recognized between Samples 70R-CC, 4.0–6.0 cm, and 66R-CC, 14.0–16.0 cm (790.285–774.982 mbsf), as an interval above the LO of D. quinqueramus (5.53 Ma) and below the FO of Ceratolithus cristatus, although the Zone NN12/NN13 (CN10b/CN10c) boundary is unreliable due to very rare discontinuous occurrences of C. cristatus.
Interval 65R-CC, 10.5–12.5 cm, through 47R-CC, 36.5–38.5 cm (769.440–682.983 mbsf), is dated to the early Pliocene as an interval from the FO of C. cristatus to the LO of Reticulofenestra pseudoumbilicus (≥7 µm) (3.82 Ma; Plate P1, figure 11). In this interval, samples from interval 65R-CC, 10.5–12.5 cm, through 61R-CC, 0.0–2.0 cm (769.440–752.165 mbsf), yielded both C. cristatus (Plate P1, figure 16) and Amaurolithus primus (Plate P1, figure 7). Zone NN15 (CN11a–NN11b) is a successive last appearance zone from the LO of A. primus (4.58 Ma) to the LO of R. pseudoumbilicus (≥7 µm) (3.82 Ma) (e.g., Young, 1998). At this site, the interval between Samples 60R-CC, 23.0–25.0 cm, and 47R-CC, 36.5–38.5 cm (746.030–682.983 mbsf), is referred to Zone NN15, although the base and top of Zone NN15 is unreliable due to discontinuous and very rare occurrences of A. primus and the possibility of reworking of R. pseudoumbilicus.
Late Pliocene Zone NN16 (CN12a–CN12c) extends from the LO of R. pseudoumbilicus (≥7 µm) (3.82 Ma) to the LO of Discoaster surculus (2.53 Ma) and was recognized between Samples 46R-CC, 28.5–30.5 cm, and 41R-CC, 19.0–21.0 cm (678.000–639.982 mbsf). In this zone, the lowermost four samples examined (Samples 46R-CC, 28.5–30.5 cm, through 43R-CC, 37.5–39.5 cm [678.000–658.982 mbsf]) were distinguished from the upper two samples (42R-CC, 49.5–51.5 cm, and 41R-CC, 19.0–21.0 cm [646.44–639.982 mbsf]) as Subzone CN12a on the basis of the LO of Discoaster tamalis (2.76 Ma) (Plate P1, figure 9). Zone NN17 (CN12c), which extends from the LO of D. surculus (2.53 Ma) to the LO of Discoaster pentaradiatus (~2.39 Ma), was not recognized in this study.
The interval between Samples 40R-CC, 28.5–30.5 cm, and 1F-CC, 19.0–21.0 cm (630.440–190.483 mbsf), is assigned to Pleistocene Zones NN18–NN21 (CN12d–CN15) as an interval above the LO of D. pentaradiatus (~2.39 Ma) to the present. Zone NN18 (CN12d) extends from the LO of D. pentaradiatus (~2.39 Ma) to the LO of Discoaster brouweri (1.93 Ma) (Plate P1, figure 15) and was recognized between Samples 40R-CC, 28.5–30.5 cm, and 38R-CC, 8.5–10.5 cm (630.440–610.035 mbsf), although the top of Zone NN18 (CN12d) is unreliable due to the rare and discontinuous occurrence of D. brouweri.
Zone NN19, which extends from the LO of D. brouweri (1.93 Ma) to the LO of Pseudoemiliania lacunosa (0.43 Ma), was recognized between Samples 37R-CC, 18.5–20.5 cm, and 22R-CC, 0.0–2.0 cm (596.780–473.515 mbsf). Zone NN19 can be divided into three subzones (CN13a, CN13b, and CN14d) following Okada and Bukry (1980). The Subzone CN13b/CN13a boundary was recognized on the basis of the FO of medium Gephyrocapsa spp. (≥3.5 µm) (1.71 Ma; Plate P1, figure 12) between Samples 36R-8, 126.5–128.5 cm, and 35R-CC, 33.5–35.5 cm (592.265–580.340 mbsf). The Subzone CN13b/CN14a boundary was defined based on the reentrance of medium-sized Gephyrocapsa oceanica (≥4.0 µm) (1.06 Ma) between Samples 29R-CC, 0.0–2.0 cm, and 27R-CC, 17.0–19.0 cm (533.810–515.380 mbsf).
The interval from Samples 21R-CC, 0.0–2.0 cm, through 14X-CC, 38.0–40.0 cm (462.100–406.390 mbsf), is assigned to Zone NN20 (CN14b) as the interval above the LO of P. lacunosa (0.43 Ma; Plate P1, figure 13) to the FO of E. huxleyi (0.29 Ma; Plate P1, figure 14). The youngest nannofossil zone (NN21 [CN15]) was recognized on the basis of the occurrence of E. huxleyi (0.29 Ma–present) in Samples 12F-CC, 3.0–5.0 cm, through 1F-CC, 19.0–21.0 cm (358.475–190.483 mbsf). The relative abundance of E. huxleyi within the total calcareous nannofossil assemblage (excluding nannoliths of F. profunda) is consistently lower than 5% in the samples from interval 12F-CC, 3.0–5.0 cm, through 2F-CC, 6.0–8.0 cm (358.475–204.175 mbsf), and ~17% in the uppermost Sample 1F-CC, 19.0–21.0 cm (190.500–190.483 mbsf) (not illustrated). Low relative abundances of E. huxleyi indicate that all samples from interval 12F-CC, 3.0–5.0 cm, through 1F-CC, 19.0–21.0 cm (358.475–190.483 mbsf), were deposited before the onset of the E. huxleyi acme (0.082–0.063 Ma; Raffi et al., 2006).
Sedimentation rate
A total of 7 reliable calcareous nannofossil events out of 12 recognized events were selected to construct the age/depth plot (Table T2; Figure F2). The sedimentation rate was very high in samples younger than 0.43 Ma (1319–610 m/My) and low in samples older than 0.43 Ma (93–35 m/My).
Table T2. Sedimentation rates based on selected calcareous nannofossil events. Download table in CSV format.
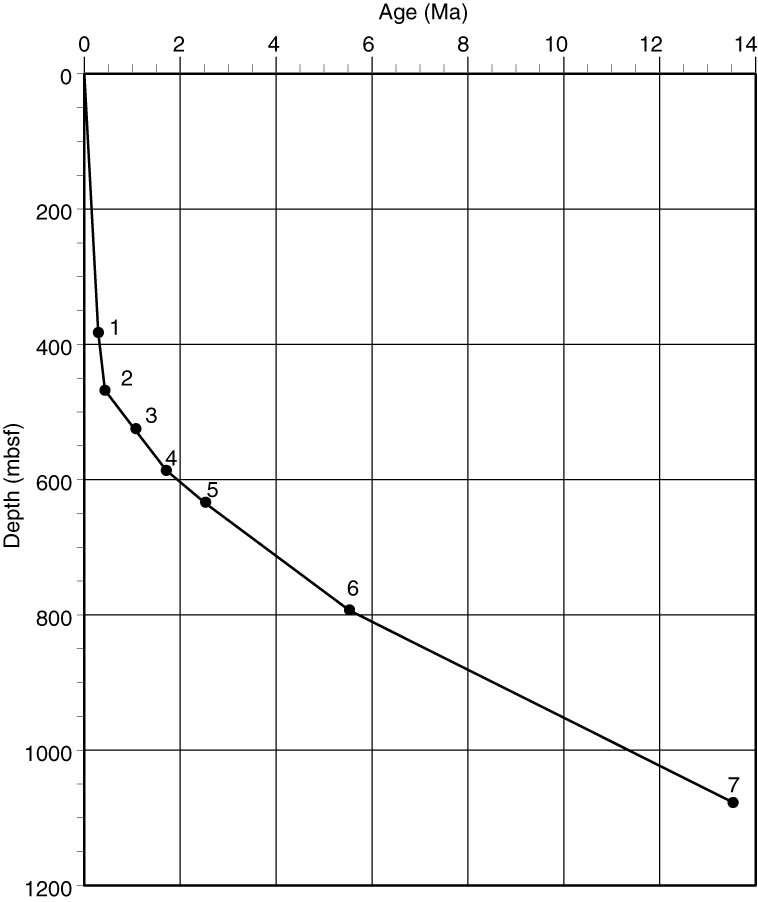
Figure F2. Age-depth plot based on selected calcareous nannofossil events.
Acknowledgments
This research used samples and data provided by the International Ocean Discovery Program (IODP), which is supported in part by the National Science Foundation. This study was supported by Grant in Aid for Scientific Research Number 17K05694 financed by the Ministry of Education, Culture, Sports, Science and Technology, Japan. We are grateful to Denise K. Kulhanek of the International Ocean Discovery Program for her critical and constructive review. We wish to express gratitude to Hisatake Okada for his suggestions on this manuscript.
References
Backman, J., Raffi, I., Rio, D., Fornaciari, E., and Pälike, H., 2012. Biozonation and biochronology of Miocene through Pleistocene calcareous nannofossils from low and middle latitudes. Newsletters on Stratigraphy, 45(3):221–244. https://doi.org/10.1127/0078-0421/2012/0022
Bown, P.R. (Ed.), 1998. Calcareous Nannofossil Biostratigraphy: Dordrecht, The Netherlands (Kluwer Academic Publishing).
Bown, P.R., and Young, J.R., 1998. Techniques. In Bown, P.R. (Ed.), Calcareous Nannofossil Biostratigraphy: Dordrecht, The Netherlands (Kluwer Academic Publishing), 16–28.
Heuer, V.B., Inagaki, F., Morono, Y., Kubo, Y., Maeda, L., Bowden, S., Cramm, M., Henkel, S., Hirose, T., Homola, K., Hoshino, T., Ijiri, A., Imachi, H., Kamiya, N., Kaneko, M., Lagostina, L., Manners, H., McClelland, H.-L., Metcalfe, K., Okutsu, N., Pan, D., Raudsepp, M.J., Sauvage, J., Schubotz, F., Spivack, A., Tonai, S., Treude, T., Tsang, M.-Y., Viehweger, B., Wang, D.T., Whitaker, E., Yamamoto, Y., and Yang, K., 2017. Expedition 370 summary. In Heuer, V.B., Inagaki, F., Morono, Y., Kubo, Y., Maeda, L., and the Expedition 370 Scientists, Temperature Limit of the Deep Biosphere off Muroto. Proceedings of the International Ocean Discovery Program, 370: College Station, TX (International Ocean Discovery Program). https://doi.org/10.14379/iodp.proc.370.101.2017
Martini, E., 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. In Farinacci, A. (Ed.), Proceedings of the Second Planktonic Conference, Roma 1970: Rome (Edizioni Tecnoscienza), 2:739–785.
Okada, H., and Bukry, D., 1980. Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973; 1975). Marine Micropaleontology, 5:321–325. https://doi.org/10.1016/0377-8398(80)90016-X
Olafsson, G., 1993. Calcareous nannofossil biostratigraphy of the Nankai Trough. In Hill, I.A., Taira, A., Firth, J.V., et al., Proceedings of the Ocean Drilling Program, Scientific Results, 131: College Station, TX (Ocean Drilling Program), 3–13. http://dx.doi.org/10.2973/odp.proc.sr.131.103.1993
Perch-Nielsen, K., 1985. Cenozoic calcareous nannofossils. In Bolli, H.M., Saunders, J.B., and Perch-Nielsen, K. (Eds.), Plankton Stratigraphy: Cambridge, United Kingdom (Cambridge University Press), 427–554.
Raffi, I., Backman, J., Fornaciari, E., Pälike, H., Rio, D., Lourens, L., and Hilgen, F., 2006. A review of calcareous nannofossil astrobiochronology encompassing the past 25 million years. Quaternary Science Reviews, 25(23–24):3113–3137. https://doi.org/10.1016/j.quascirev.2006.07.007
Shipboard Scientific Party, 1991. Site 808. In Taira, A., Hill, I., Firth, J.V., et al., Proceedings of the Ocean Drilling Program, Initial Reports, 131: College Station, TX (Ocean Drilling Program), 71–269. http://dx.doi.org/10.2973/odp.proc.ir.131.106.1991
Shipboard Scientific Party, 2001. Site 1174. In Moore, G.F., Taira, A., Klaus, A., et al., Proceedings of the Ocean Drilling Program, Initial Reports, 190: College Station, TX (Ocean Drilling Program), 1–149. http://dx.doi.org/10.2973/odp.proc.ir.190.105.2001
Young, J.R., 1998. Neogene. In Bown, P.R. (Ed.), Calcareous Nannofossil Biostratigraphy: Dordrecht, The Netherlands (Kluwer Academic Publishing), 225–265.
Appendix
Systematic paleontology
Bibliographic references for these taxa can be found in Perch-Nielsen (1985) and Bown (1998).
Amaurolithus delicatus Gartner and Bukry, 1975
Amaurolithus primus (Bukry and Percival, 1971) Gartner and Bukry, 1975
Amaurolithus tricorniculatus (Gartner, 1967) Gartner and Bukry, 1975
Braarudosphaera bigelowii (Gran and Braarud, 1935) Deflandre, 1947
Calcidiscus leptoporus (Murray and Blackman, 1898) Loeblich and Tappan, 1978
Calcidiscus tropicus Kamptner, 1956 sensu Gartner, 1992
Calcidiscus macintyrei (Bukry and Bramlette, 1969) Loeblich and Tappan, 1978
Catinaster calyculus Martini and Bramlette, 1963
Catinaster coalitus Martini and Bramlette 1963
Ceratolithus armatus Müller, 1974
Ceratolithus cristatus Kamptner, 1950
Coccolithus pelagicus (Wallich, 1871) Schiller, 1930
Coccolithus miopelagicus Bukry, 1971
Cyclicargolithus floridanus (Roth and Hay in Hay et al., 1967) Bukry, 1971
Discoaster asymmetricus Gartner, 1969
Discoaster berggrenii Bukry, 1971
Discoaster bollii Martini and Bramlette, 1963
Discoaster calcaris Gartner 1967
Discoaster deflandrei Bramlette and Riedel, 1954
Discoaster exilis Martini and Bramlette, 1963
Discoaster kugleri Martini and Bramlette, 1963
Discoaster pentaradiatus Tan, 1927
Discoaster quadramus Bukry, 1973
Discoaster quinqueramus Gartner 1969
Discoaster surculus Martini and Bramlette, 1963
Discoaster tamalis Kamptner, 1967
Discoaster triradiatus Tan, 1927
Discoaster variabilis Martini and Bramlette, 1963
Emiliania huxleyi (Lohmann, 1902) Hay and Mohler in Hay et al., 1967
Florisphaera profunda Okada and Honjo, 1973
Gephyrocapsa ericsonii McIntyre and Bé, 1967
Gephyrocapsa oceanica Kamptner, 1943
Helicosphaera carteri (Wallich, 1877) Kamptner, 1954
Helicosphaera granulata (Bukry and Percival, 1971) Jafar and Martini, 1975
Helicosphaera princei da Gama and Varol, 2013
Helicosphaera sellii (Bukry and Bramlette, 1969) Jafar and Martini, 1975
Helicosphaera wallichii (Lohmann, 1902) Okada and McIntyre, 1977
Orthorhabdus rugosus (Bramlette and Wilcoxon, 1967) Young and Bown, 2014
Pseudoemiliania lacunosa (Kamptner, 1963) Gartner, 1969
Pseudoemiliania ovata (Bukry, 1973) Young, 1998
Reticulofenestra asanoi Sato and Takayama, 1992
Reticulofenestra bisecta (Hay, Mohler, and Wade, 1966) Roth, 1970
Reticulofenestra haqii Backman, 1978
Reticulofenestra minuta Roth, 1970
Reticulofenestra minutula (Gartner, 1967) Haq and Berggren, 1978
Reticulofenestra pseudoumbilicus (Gartner, 1967) Gartner, 1969
Reticulofenestra productella (Bukry, 1975) Gallagher, 1989
Sphenolithus abies Deflandre in Deflandre and Fert, 1954
Sphenolithus heteromorphus Deflandre, 1953
Sphenolithus moriformis (Bronnimann and Stradner, 1960) Bramlette and Wilcoxon, 1967
Umbellosphaera irregularis Paasche in Markali and Paasche, 1955
Umbilicosphaera rotula (Kamptner, 1956) Varol, 1982
Umbilicosphaera sibogae (Weber van Bosse, 1901) Gaarder, 1970
2 Centre for Advanced Marine Core Research, Kochi University, Japan. hagino@kochi-u.ac.jp
3 Expedition 370 Scientists’ addresses.
This work is distributed under the Creative Commons Attribution 4.0 International (CC BY 4.0) license. 