Li, C.-F., Lin, J., Kulhanek, D.K., and the Expedition 349 Scientists
Proceedings of the International Ocean Discovery Program Volume 349
publications.iodp.org

doi:10.14379/iodp.proc.349.201.2016
Data report: Oligocene foraminifers and stable isotopes from IODP Hole U1435A1
Qianyu Li,2, 3 Xinrong Cheng,2 Jingshuang Chen,2 and Jianming Xu2
Keywords: International Ocean Discovery Program, IODP, JOIDES Resolution, Expedition 349, Site U1435, Hole U1435A, South China Sea, Oligocene, foraminifer, stable isotopes
MS 349-201: Received 1 February 2016 · Accepted 18 July 2016 · Published 25 August 2016
Abstract
The foraminifer assemblage in samples from the Oligocene section in International Ocean Discovery Program Hole U1435A contains abundant specimens of planktonic foraminifers and frequent benthic foraminifers. Planktonic species indicate Zones P18 and P19 deposition in the early Oligocene, whereas benthic species imply outer-shelf to slope settings. Both planktonic and benthic δ18O show progressive enrichment upsection.
Introduction
International Ocean Discovery Program (IODP) Expedition 349 Site U1435 is located on a structural high at the transition between the extended continental crust and the oceanic crust in the northern South China Sea (SCS), at a water depth of 3757 m (Figure F1). As the shallowest of five sites drilled during the expedition, Site U1435 was designed to recover an older (possibly Mesozoic) sequence that may have formed the basement before break up and spreading of the SCS. Hole U1435A was drilled to 300 m below seafloor (mbsf) and recovered three units. Unit I (0–77.65 mbsf) is a sequence of Oligocene–Pleistocene greenish gray nannofossil-rich clay and clayey nannofossil ooze and is divided into Subunits IA and IB. Subunit IA (0–36.04 mbsf) is Miocene to Pleistocene in age and consists of manganese nodules underlain by clayey nannofossil ooze. Subunit IB (36.04–77.65 mbsf) is Oligocene in age and consists of mostly greenish gray nannofossil-rich clay and lesser quantities of greenish gray clay, which is heavily bioturbated with trace fossils of the Nereites ichnofacies. Unit II (77.65–275.54 mbsf) is a 197.89 m thick sequence of pre-Oligocene thick-bedded and mostly medium-grained dark gray silty sandstone, with very little carbonate and minor sandy siltstone and conglomerate and burrows typical of the Cruziana ichnofacies indicative of shallow-marine conditions. Unit III (275.54–300.00 mbsf) is a 24.46 m thick sequence of dark gray silty sandstone, silty mudstone, and minor conglomerate (see the Site U1435 chapter [Li et al., 2015b]).
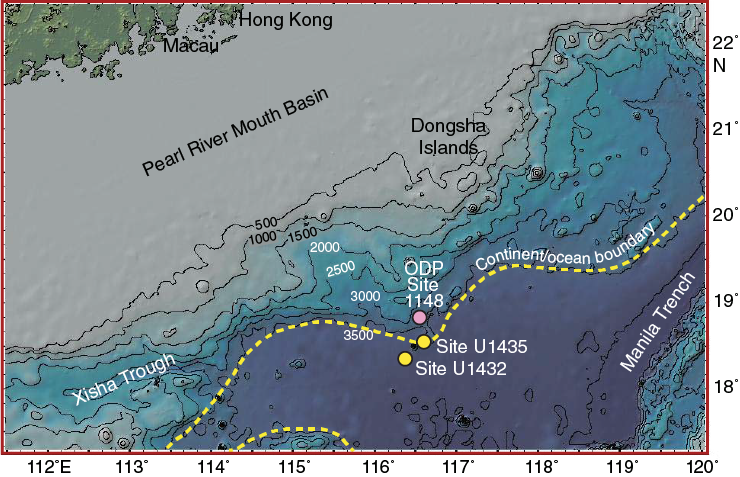
Figure F1. Bathymetry and site locations.
We investigated planktonic and benthic foraminifers and measured their stable isotopes in samples from Subunit IB of Hole U1435A. The primary objective of this study was to record the distribution patterns of foraminifers and to provide information for interpreting Oligocene environmental changes during the early history of the SCS.
Materials and methods
Subunit IB of Hole U1435A is represented by 21 m of core with 51% recovery (see the Site U1435 chapter [Li et al., 2015b]). The lower section (especially between 58.6–71.6 mbsf) contains several dolomite-rich layers. Preliminary shipboard paleontological investigation indicated that the section represents full marine deposition during the Oligocene, likely immediately after the breakup of the SCS (see the Site U1435 chapter [Li et al., 2015b]).
For this study, we collected 43 samples (20 cm3) from split sections and 4 samples from core catchers over Subunit IB, yielding a sampling resolution of mostly 30–60 cm. Samples were oven-dried at 60°C, weighed, and then washed through a 63 µm sieve. Dry residues were weighed again for the purpose of calculating the >63 μm coarse fraction. Planktonic and benthic foraminifers in the dry residues were analyzed under a binocular microscope. Relative abundances of species or species groups were estimated using the following categories: D = dominant (>30% of the assemblage), A = abundant (10%−30%), F = few (5%−10%), R = rare (1%−5%), and P = present (<1%). We referred to Bolli and Saunders (1985) and Kennett and Srinivasan (1983) for identification of planktonic foraminifer species, to Kuhnt et al. (2002) and Ortiz and Thomas (2006) for benthic species, and to Wade et al. (2011) for zonation, and to Gradstein et al. (2012) for age of datums.
Specimens of planktonic species Chiloguembelina cubensis (40–60 specimens; 100–150 µm), Cassigerinella chipolensis (40–60 specimens; 100–150 µm), Globigerina ciperoensis (20–30 specimens; 150–250 µm), and Globoquadrina venezuelana (5–10 specimens; 300–400 µm), and benthic species Cibicidoides spp. (3–5 specimens; 400–450 µm) were picked for measuring stable isotopes. The picked specimens were washed with ethanol (~99.7%) in an ultrasonic bath, dried in an oven at 60°C, and reacted with orthophosphoric acid in an automated carbonate device (Kiel III) at 70°C to generate CO2, which was then transferred to a Finnigan MAT252 mass spectrometer for measuring stable isotopes. Because of their small test size, both C. cubensis and C. chipolensis often required repeat analyses on a new set of specimens for better results. Measurement precision was regularly checked with a Chinese national carbonate standard GBW04405; the standard deviation is 0.07‰ for δ18O and 0.05‰ for δ13C. Finally, an NBS19 standard was used to convert the results to the international Peedee belemnite (PDB) scale. All analyses and measurements were performed in the State Key Laboratory of Marine Geology of Tongji University.
Results
The original planktonic foraminifer census data plus notes on the benthic species groups are shown in Table T1. In general, both planktonic and benthic foraminifers are extremely rare in the lowermost Sample 349-U1435A-9R-2, 125–129 cm (77.55–77.59 mbsf), which is characterized by fine sand similar to the lithology in underlying Unit II. Planktonic and benthic foraminifer abundance increases rapidly from Sample 9R-2, 60–64 cm (76.90–76.94 mbsf), uphole, with group abundance from common to abundant (40%–70% or more) over the rest of the studied interval. Planktonic/benthic ratio (P/B) also quickly reached 70:30 to 90:10 and stayed high with values of 80–90:20–10 except in samples between 58.6 and 71.6 mbsf, which are influenced by dolomite sand. Preservation is mostly good with no sign of obvious dissolution. The foraminifer assemblage is dominated by small specimens and only in several samples were some large tests observed, such as 8R-5, 11–15 cm (70.90–70.94 mbsf), as well as most samples from shallower than 58 mbsf (Table T1).
Table T1. Foraminfer census data. Download table in .csv format.
Planktonic foraminifers and zones
The most common planktonic foraminifer species include Chiloguembelina cubensis, Cassigerinella chipolensis, Tenuitella spp., and Pseudohastigerina naguewichiensis, all with a small test size of 100–150 µm. Species with a medium test size of ~200–250 µm, such as Globigerina praebulloides (and allied forms), Globigerina ciperoensis, Globorotaloides suteri, Catapsydrax spp. and Dentoglobigerina spp., are frequent, whereas those with a large test size of 300 µm or larger, including Turborotalia ampliapertura and Globoquadrina spp., are few or rare (Table T1; Figure F2).
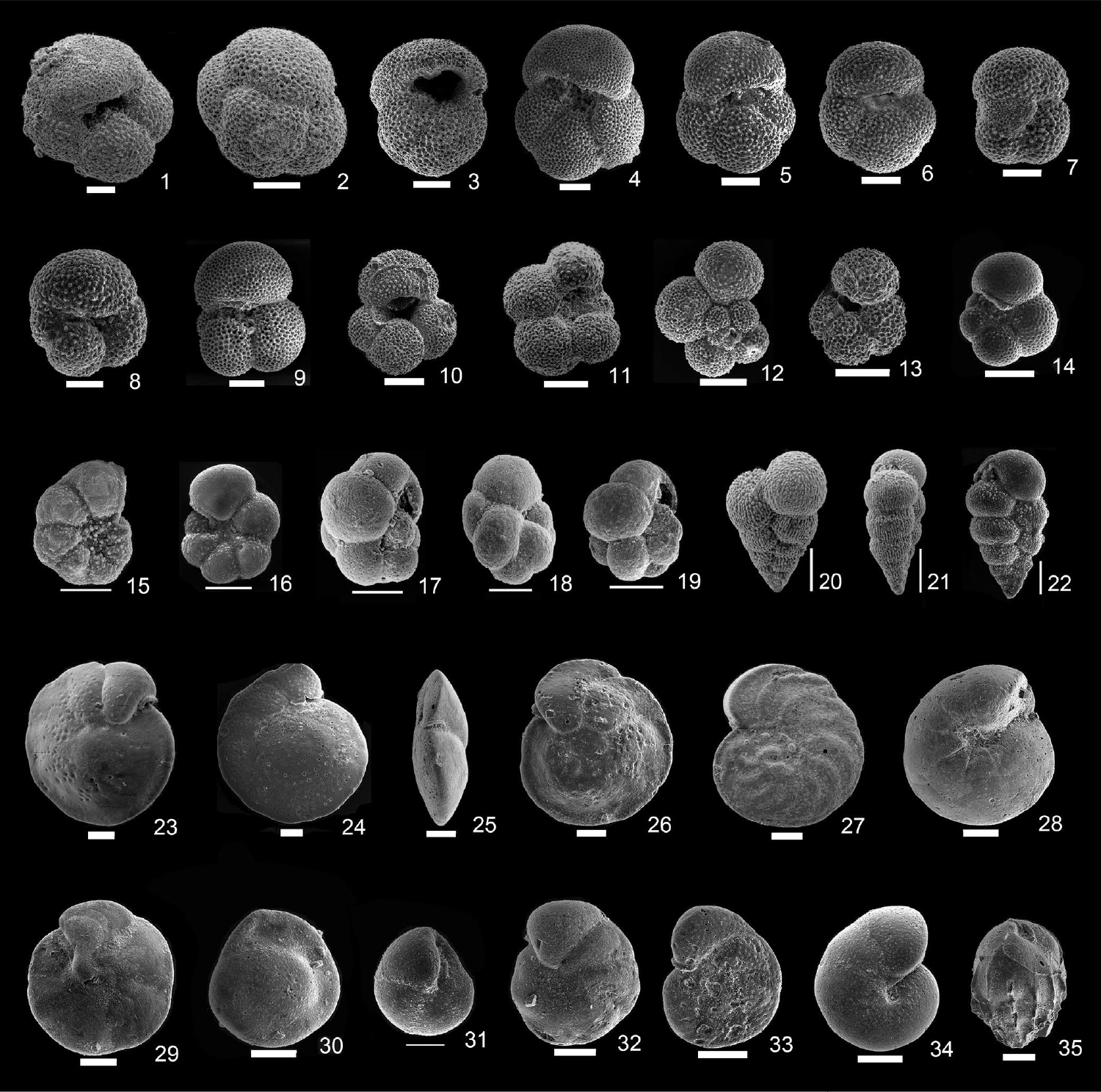
Figure F2. Selected foraminifers.
These species represent a typical early Oligocene assemblage (Bolli and Saunders, 1985; Wade et al., 2011). Specifically, the top and base of P. naguewichiensis occurs, respectively, in Samples 349-U1435A-6R-1, 110–114 cm (46.80–46.84 mbsf), and 9R-2, 60–64 cm (76.90–76.94 mbsf), indicating an age of 32.10 Ma and older. The latter sample also contains the first occurrence of C. chipolensis (base at 33.89 Ma). The co-occurrence of P. naguewichiensis and C. chipolensis between 46.80 and 76.94 mbsf indicates the early Oligocene part of Zone P18, from <33.89 to 32.10 Ma (Figure F3). Therefore, the base of Subunit IB in Hole U1435A is earliest Oligocene in age. The absence of late Eocene species such as Pseudohastigerina micra and Hantkenina spp. (both with tops at 33.89 Ma) from these samples further supports this observation. However, whether Sample 9R-2, 60–64 cm (76.90–76.94 mbsf), marks the real base of C. chipolensis or a younger pseudobase needs further scrutiny because the sample is the base of the recovered marine section. Similarly, T. ampliapertura (base at ~35 Ma) first occurs in Sample 9R-1, 60–64 cm (75.40–75.44 mbsf), rather than the lowermost sample within the marine section. The observed base may not represent its real base because the underlying samples are dominated by small species instead of T. ampliapertura-like large tests. Thus, we assign the base of Subunit IB as representing deposition younger than 33.89 Ma.
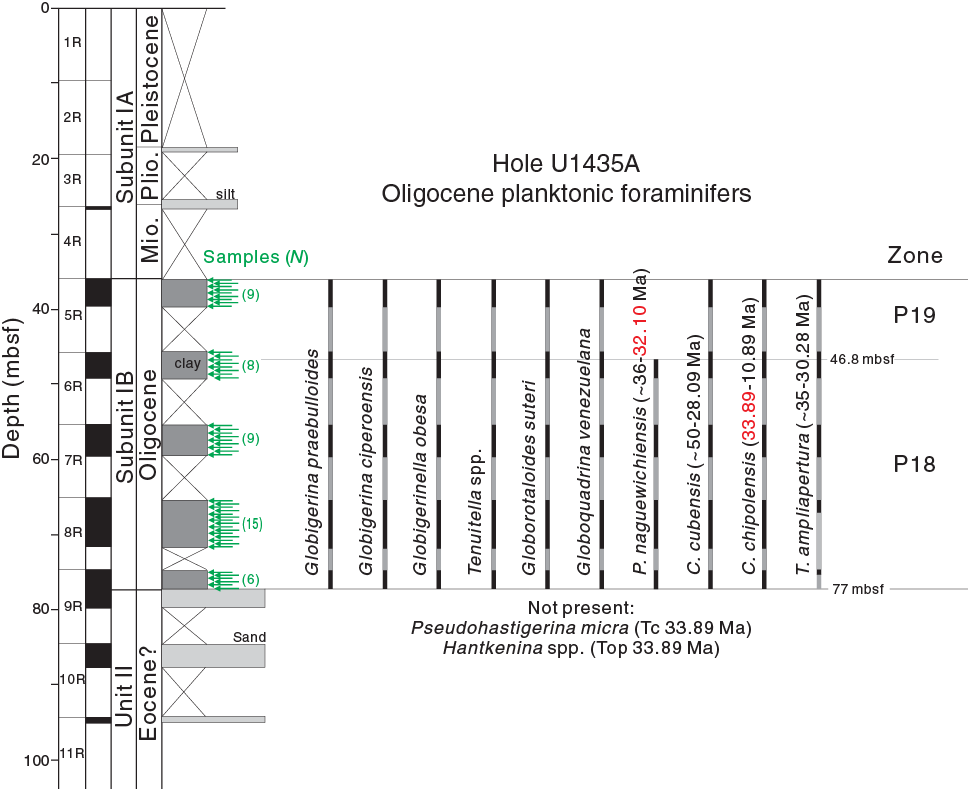
Figure F3. Foraminifer species ranges.
For the upper part of Subunit IB, T. ampliapertura (top at 30.28 Ma) is present, but Paragloborotalia opima opima (base at 30.72 Ma) is not, although its allied form Paragloborotalia opima nana and their transitional types are found between Samples 5R-3, 52–56 cm (39.30–39.34 mbsf), and 5R-1, 70–74 cm (36.70–36.74 mbsf). Therefore, the top of Subunit IB is placed in the middle part of Zone P19, or older than 30.72 Ma (Figure F3).
Benthic foraminifers
The benthic foraminifer assemblage consists of a very diverse population of species commonly found in upper- to middle-slope environments (e.g., van Morkhoven et al., 1986). Calcareous hyaline species are dominant, including many species of lagenids (Dentalina, Chrysalogonium, Fissurina, Stilostomella, Guttulina, and Lenticulina), buliminids (Bolivina, Bulimina, Uvigerina, Trifarina, Pleurostomella, and Plectofrondicularia), and planispiral and trochospiral rotaliids (Cassidulina, Globocassidulina, Pullenia, Melonis, Gyroidina, Cibicidoides, Planulina, Oridorsalis, and Epistominella) (Figure F1). Agglutinated forms, represented by Textularia, Bolivinopsis, Karreriella, and Vulvulina, are rare, whereas porcelaneous species are sporadic or absent. Many species are identical to those described in samples from Ocean Drilling Program Site 1148 by Kuhnt et al. (2002). Noteworthy is that most specimens are medium to small in size, and only in selected samples (i.e., Samples 349-U1435A-9R-1, 60–64 cm [75.40–75.44 mbsf], 8R-5, 11–15 cm [70.90–70.94 mbsf], and 7R-2, 25–29 cm [57.15–57.19 mbsf]), are large benthic tests frequent in number. Some characteristic calcareous species are shown in Figure F2.
Compared to the planktonic foraminifers, benthic specimens are always less abundant, with percentages decreasing rapidly from 30% in Sample 9R-2, 60–64 cm (76.90–76.94 mbsf) to ~10% or less in samples immediately above (Table T1). Although planktonic species are often represented by many individual specimens, most benthic species occur only with one or several specimens. Reworked specimens are largely filled or replaced with pyrite and show abrasive features and different colors. Reworked specimens are always rare.
Unlike the calcareous species–dominated assemblage in Hole U1435A, the benthic assemblage found in the lower Oligocene section of Site 1148, which is located ~50 km north of Site U1435 (Figure F1), is characterized by large, elongate, sometimes branching agglutinated forms (Kuhnt et al., 2002; Zhao et al., 2009). Instead, their late Oligocene–early Miocene assemblage contains abundant calcareous hyaline tests, similar to the assemblage found in the lower Oligocene Subunit IB of Hole U1435A.
The benthic foraminifer assemblage found in Subunit IB in Hole U1435A indicates upper- to middle-slope settings. The transition to slope from outer shelf is represented by Sample 9R-2, 60–64 cm (76.90–76.94 mbsf), with a benthic proportion of ~30% from the base of the subunit. This sample also contains more pyrite grains and small benthic forms with frequent Cibicides, Textularia, Bigenerina, Karreriella, Trifarina, Cassidulina, Bolivina, Uvigerina, Fissurina, and Denticulina, which are characteristic of the benthic assemblage from ~150–500 m present-day water depths (Wang et al., 1985; Szarek et al., 2006, 2009).
Stable isotopes
Stable isotope results are shown in Table T2 and plotted in Figure F4. The results show that δ18O in Hole U1435A samples varies between –5‰ and –0.9‰ for planktonic species and between –3‰ and 1.5‰ for benthic species, whereas δ13C varies between –2.76‰ and 1.4‰ for planktonic species and between –1‰ and 0.9‰ for benthic species. The lightest δ13C value (–2.76‰), recorded for C. chipolensis in Sample 349-U1435A-5R-1, 130–134 cm (37.30–37.34 mbsf), may be a measuring error because other planktonic species from the same sample register –1‰ or heavier δ13C values.
Table T2. Foraminifer isotopic data. Download table in .csv format.
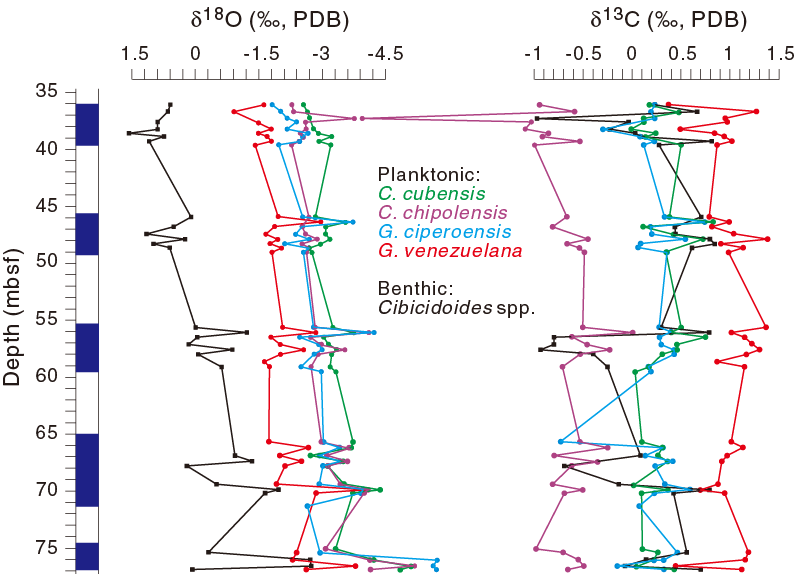
Figure F4. Foraminfer stable isotope variations.
Both benthic and planktonic δ18O show an enriched trend, and variations between the two groups become slightly enlarged upsection. The δ18O values are heaviest for Cibicidoides spp., followed by G. venezuelana, G. ciperoensis, C. chipolensis, and C. cubensis, although the δ18O differences between G. ciperoensis and C. chipolensis are minor. Our results support the δ18O patterns of these species as reported in early studies (e.g., Keller, 1985; Pearson et al., 1997).
Wider variations are displayed in the δ13C records. For the benthic δ13C, three negative shifts can be observed at about 69, 57, and 37 mbsf, respectively (Figure F4). Planktonic δ13C appears similar but shows minor positive shifts at 69 and 57 mbsf instead of the pronounced negative shifts seen in the benthic record. Both benthic and planktonic δ13C records lack distinct heavy values characteristic of the global δ13C maximum across the Eocene/Oligocene boundary (Zachos et al., 2001). The absence of such a positive δ13C shift from the record is further evidence that the Subunit IB marine section in Hole U1435A was deposited after the boundary event, or younger than 33.89 Ma.
Summary
Planktonic foraminifer Zones P18 and P19 of early Oligocene age are recognized in samples from Subunit IB of IODP Hole U1435A. The sediment section is dated to between <33.89 and >30.72 Ma based on the occurrence of Cassidulinella chipolensis and T. ampliapertura and the absence of P. opima opima. The Zone P18/P19 boundary lies close to 46.8 mbsf, as evidenced by the top of P. naguewichiensis.
The benthic foraminifer assemblage is dominated by calcareous hyaline species that live in upper- to middle-slope settings. Shallower water depths of 150–500 m are indicated by the benthic assemblage from the lowermost part of the section
Benthic and planktonic δ18O values increase progressively upsection without any obvious sudden shifts. Their δ13C variations are larger, but larger positive swings matching the Oligocene δ13C maxima are lacking.
Acknowledgments
This research used samples and data provided by the International Ocean Discovery Program (IODP). Expedition 349 scientists and technicians helped with sampling and provided logistical support. Funding for this research was provided by IODP China Secretariat and the National Natural Science Foundation of China (grant number 91228203). Wolfgang Kuhnt and Denise Kulhanek are thanked for valuable comments and careful editorial corrections that greatly improved this contribution.
References
Bolli, H.M., and Saunders, J.B., 1985. Oligocene to Holocene low latitude planktic foraminifera. In Bolli, H.M., Saunders, J.B., and Perch-Nielsen, K. (Eds.), Plankton Stratigraphy (Volume 1): Planktic Foraminifera, Calcareous Nannofossils and Calpionellids: Cambridge, United Kingdom (Cambridge University Press), 155–262.
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. (Eds.), 2012. The Geological Time Scale 2012: Amsterdam (Elsevier).
Keller, G., 1985. Depth stratification of planktonic foraminifers in the Miocene ocean. In Kennett, J.P. (Ed.), The Miocene Ocean: Paleoceanography and Biogeography. Geological Society of America Memoirs, 163:177–196. http://dx.doi.org/10.1130/MEM163-p177
Kennett, J.P., and Srinivasan, M.S., 1983. Neogene Planktonic Foraminifera: A Phylogenetic Atlas: Stroudsburg, PA (Hutchinson Ross).
Kuhnt, W., Holbourn, A., and Zhao, Q., 2002. The early history of the South China Sea: evolution of Oligocene-Miocene deep water environments. Revue de Micropaléontologie, 45:99–159.
Li, C.-F., Lin, J., Kulhanek, D.K., Williams, T., Bao, R., Briais, A., Brown, E.A., Chen, Y., Clift, P.D., Colwell, F.S., Dadd, K.A., Ding, W., Hernández-Almeida, I., Huang, X.-L., Hyun, S., Jiang, T., Koppers, A.A.P., Li, Q., Liu, C., Liu, Q., Liu, Z., Nagai, R.H., Peleo-Alampay, A., Su, X., Sun, Z., Tejada, M.L.G., Trinh, H.S., Yeh, Y.-C., Zhang, C., Zhang, F., Zhang, G.-L., and Zhao, X., 2015a. Expedition 349 summary. In Li, C.-F., Lin, J., Kulhanek, D.K., and the Expedition 349 Scientists, South China Sea Tectonics. Proceedings of the International Ocean Discovery Program, 349: College Station, TX (International Ocean Discovery Program). http://dx.doi.org/10.14379/iodp.proc.349.101.2015
Li, C.-F., Lin, J., Kulhanek, D.K., Williams, T., Bao, R., Briais, A., Brown, E.A., Chen, Y., Clift, P.D., Colwell, F.S., Dadd, K.A., Ding, W., Hernández-Almeida, I., Huang, X.-L., Hyun, S., Jiang, T., Koppers, A.A.P., Li, Q., Liu, C., Liu, Q., Liu, Z., Nagai, R.H., Peleo-Alampay, A., Su, X., Sun, Z., Tejada, M.L.G., Trinh, H.S., Yeh, Y.-C., Zhang, C., Zhang, F., Zhang, G.-L., and Zhao, X., 2015b. Site U1435. In Li, C.-F., Lin, J., Kulhanek, D.K., and the Expedition 349 Scientists, South China Sea Tectonics. Proceedings of the International Ocean Discovery Program, 349: College Station, TX (International Ocean Discovery Program). http://dx.doi.org/10.14379/iodp.proc.349.107.2015
Ortiz, S., and Thomas, E., 2006. Lower-middle Eocene benthic foraminifera from the Fortuna Section (Betic Cordillera, southeastern Spain). Micropaleonotology, 52(2):97–150. http://dx.doi.org/10.2113/gsmicropal.52.2.97
Pearson, P.N., and Shackleton, N.J., 1995. Neogene multispecies planktonic foraminifer stable isotope record, Site 871, Limalok Guyot. In Haggerty, J.A., Premoli Silva, I., Rack, F., and McNutt, M.K. (Eds.), Proceedings of the Ocean Drilling Program, Scientific Results, 144: College Station, TX (Ocean Drilling Program), 401–410. http://dx.doi.org/10.2973/odp.proc.sr.144.054.1995
Pearson, P.N., Shackleton, N.J., Weedon, G.P., and Hall, M.A., 1997. Multispecies planktonic foraminifer stable isotope stratigraphy through Oligocene/Miocene boundary climatic cycles, Site 926. In Shackleton, N.J., Curry, W.B., Richter, C., and Bralower, T.J. (Eds.), Proceedings of the Ocean Drilling Program, Scientific Results, 154: College Station, TX (Ocean Drilling Program), 441–449. http://dx.doi.org/10.2973/odp.proc.sr.154.118.1997
Szarek, R., Kuhnt, W., Kawamura, H., and Kitazato, H., 2006. Distribution of recent benthic foraminifera on the Sunda Shelf (South China Sea). Marine Micropaleontology, 61(4):171–195. http://dx.doi.org/10.1016/j.marmicro.2006.06.005
Szarek, R., Kuhnt, W., Kawamura, H., and Nishi, H. 2009. Distribution of recent benthic foraminifera along continental slope of the Sunda Shelf (South China Sea). Marine Micropaleontology, 71(1–2):41–59. http://dx.doi.org/10.1016/j.marmicro.2009.01.004
van Morkhoven, F.P.C.M., Berggren, W.A., Edwards, A.S., and Oertli, H.J., 1986. Cenozoic cosmopolitan deep-water benthic foraminifera. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 11.
Wade, B.S., Pearson, P.N., Berggren, W.A., and Pälike, H., 2011. Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth-Science Reviews, 104(1–3):111–142. http://dx.doi.org/10.1016/j.earscirev.2010.09.003
Wang, P.X., Min, Q.B., and Bian, Y.H., 1985. Foraminiferal biofacies in the northern continental shelf of the South China Sea. In Wang, P.X. (Ed.), Marine Micropaleontology of China: Beijing (China Ocean Press), 151–175.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K., 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science, 292(5517):686–693. http://dx.doi.org/10.1126/science.1059412
Zhao, Q., Li, Q., and Jian, Z., 2009. Deep waters and oceanic connection. In Wang, P., and Li, Q. (Eds.), Developments in Paleoenvironmental Research (Volume 13): The South China Sea: Paleoceanography and Sedimentology: Amsterdam (Springer), 395–437. http://dx.doi.org/10.1007/978-1-4020-9745-4_6
1Li, Q., Cheng, X., Chen, J., and Xu, J., 2016. Data report: Oligocene foraminifers and stable isotopes from IODP Hole U1435A. In Li, C.-F., Lin, J., Kulhanek, D.K., and the Expedition 349 Scientists, South China Sea Tectonics. Proceedings of the International Ocean Discovery Program, 349: College Station, TX (International Ocean Discovery Program). http://dx.doi.org/10.14379/iodp.proc.349.201.2016
2State Key Laboratory of Marine Geology, School of Ocean and Earth Sciences, Tongji University, Shanghai 200092, China. Correspondence author: qli01@tongji.edu.cn
3Also at: School of Earth and Environmental Sciences, University of Adelaide, Adelaide SA 5005, Australia.